
Part:BBa_K3781001
SARS-CoV-2 spike protein receptor binding domain, MocloMania B3_B4

This part codes for the receptor binding domain of the SARS-CoV-2 spike protein. These spike proteins are part of multimeric protein structures called spikes that project outwards from the virus envelope’s outer surface.[1] As one of the four viral structural proteins it is essential for the recognition, binding and entry of the virus into the host’s cells.[2] This is mediated by the receptor binding domain, located in the S1 region of the spike protein, due to its specific binding to the angiotensin-converting enzyme 2 (ACE2) located on the outer membrane of many mammalian and bird cells.[3] This binding results in a TMPRSS2 mediated proteolytic cleavage of the spike protein S2 region, exposing a viral fusion peptide and thus enabling virus entry into the host cells.[4]
size 25.1 kDa
function coding sequence
cloning position B3_B4
plasmid backbone pAGM1287
Data
We were able to successfully clone this basic part into its respective L0 plasmid backbone and to confirm the integrity of the L0 construct via restriction digest and gel electrophoresis, see Figure 1. Furthermore, we were able to include it into a variety of L1 constructs, proving its correct adaptation towards MoClo assembly, see Figure 2.
-
 Figure 1 | Test digest of L0_RBD_B3_B4 using BsaI
Figure 1 | Test digest of L0_RBD_B3_B4 using BsaI
1 | pAGM1287 | 2247 + 598 bp
2 | L0_RBD_B3_B4 | 2247 + 675 bp
L | Thermofischer GeneRuler Plus Ladder [bp] -
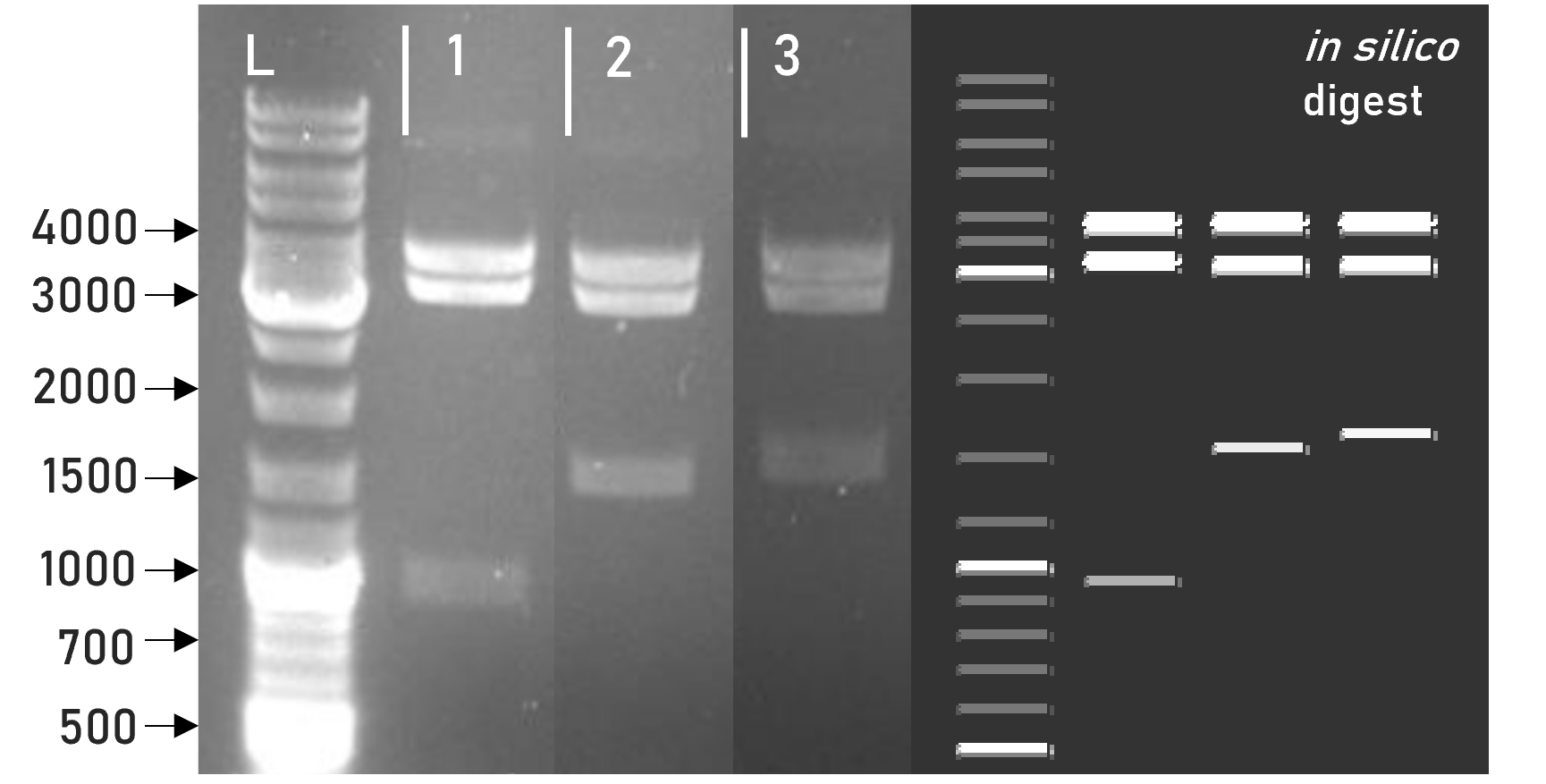 Figure 2 | Test digest of L1 constructs using HindIII
Figure 2 | Test digest of L1 constructs using HindIII
1 | L1_sAP_RBD_3xHA | 3792 + 3198 + 981 bp
2 | L1_sAP_RBD_GST | 3792 + 3198 + 1563 bp
3 | L1_sAP_mCerulean_GST | 3792 + 3198 + 1656 bp
L | Thermofischer GeneRuler Plus Ladder [bp]


Since our project's objective was to test the efficiency of our MocloMania expression system by expressing and purifying recombinant SARS-CoV-2 receptor binding domain, this basic part was incorporated into a vast variety of different L1 constructs. For this, L0_RBD_B3_B4 was usually fused to the N-terminal sAP secretion tag and a variable C-terminal purification or detection tag. Most of these L1 constructs have been successfully transfected into Leishmania, resulting in recombinant protein expression that could be observed via immunostaining on western blot, see Figures 3-5.
-
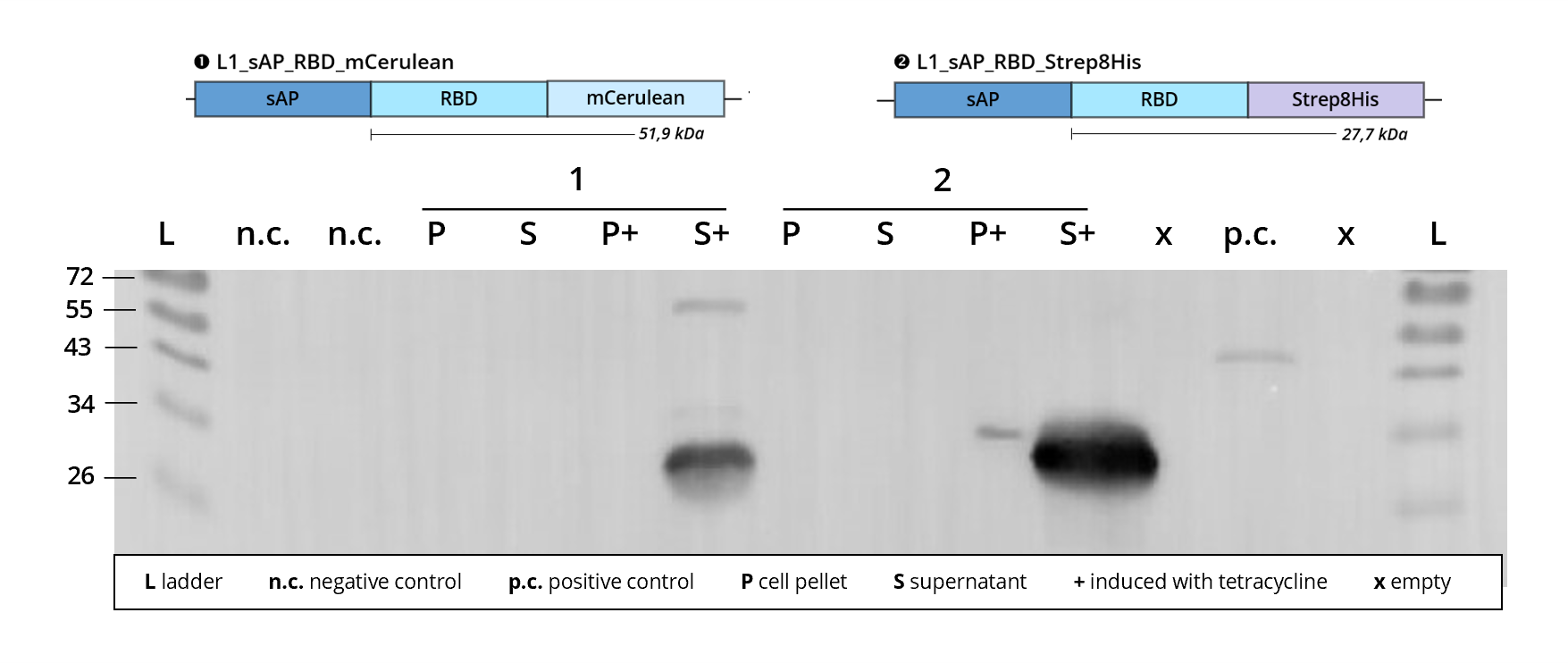 Figure 3 | Immunoblot of L1 transfected Leishmania | stained against RBD
Figure 3 | Immunoblot of L1 transfected Leishmania | stained against RBD
1 | L1_sAP_RBD_mCerulean | 51.9 kDa
2 | L1_sAP_RBD_Strep8His | 27.7 kDa
n.c. | negative control | untransfected Leishmania culture
p.c. | RBD-GFP | 54 kDa
L | Thermofischer PageRuler Protein Ladder [kDa]
1. AB | ms anti-RBD | 1:2,000
2. AB | rb anti-ms HRP | 1:10,000 -
 Figure 4 | Immunoblot of L1 transfected Leishmania | stained against RBD
Figure 4 | Immunoblot of L1 transfected Leishmania | stained against RBD
1 | L1_sAP_RBD_mCerulean | 51.9 kDa
2 | L1_sAP_RBD_mCerulean_Strep8His | 54.3 kDa
3 | L1_sAP_RBD_mCerulean_GST | 78.6 kDa
4 | L1_3xHA_RBD_mCerulean | 55.7 kDa
n.c. | negative control | Leishmania culture
transfected with empty L1 expression vector p.c. | RBD-GFP | 54 kDa
L | Thermofischer PageRuler Protein Ladder [kDa]
1. AB | ms anti-RBD
2. AB | rb anti-ms HRP -
 Figure 5 | Immunoblot of L1 transfected Leishmania cell cultures | stained against RBD
Figure 5 | Immunoblot of L1 transfected Leishmania cell cultures | stained against RBD
1 | L1_sAP_RBD_Strep8His | 27.7 kDa
2 | L1_sAP_Myc_RBD_Strep8His | 30.2 kDa
3 | L1_sAP_RBD_GST | 51.9 kDa
n.c. | negative control | untransfected Leishmania culture
L | Thermofischer PageRuler Protein Ladder [kDa]
1. AB | ms anti-RBD | 1:2,000
2. AB | rb anti-ms HRP | 1:10,000



In all of the blots shown above, the constructs carrying L0_RBD_B3_B4 were successfully stained with the help of anti-RBD antibodies. This verifies the correct recombinant expression of the underlying basic part sequence. In order to further analyse the quality of the expressed receptor binding domain, analysis blots were follwed by purification procedures to harvest produced RBD for downstream activity assays. So far, only L1_sAP_RBD_GST showed significant purification results, see Figure 6.
-
 Figure 6 | Immunoblot | L1_sAP_RBD_GST | after GST purification
Figure 6 | Immunoblot | L1_sAP_RBD_GST | after GST purification
1. AB | gt anti-GST | 1:10,000
2. AB | rb anti-goat HRP | 1:2,000
I Input | FT Flowthrough | W Wash | E Eluate | E.T Eluate, TCA precipitated
L.C | cell lysate of untransfected negative control
S.C | supernatant of untransfected negative control
S.A | supernatant after pelleting of ammonium precipitation
P.A | resuspended pellet of ammonium sulfate precipitation
GST | GST positive control | 26 kDa
RBD | RBD-GFP positive control | 52 kDa
L | Thermo Scientific PageRuler Prestained Protein Ladder [kDa]

Here, the RBD_GST fusion protein was stained with anti-GST antibodies. This doesn't only visualize RBD_GST (expected around 52 kDa), but also singular GST protein expected at around 26 kDa. In order to read more on the interpretation of this band pattern as well as on the poor purification efficiency, please consult the L0_GST_B5 part page.
Despite very low purification yield, the samples resulting from GST-purification, along with other constructs carrying L0_RBD_B3_B4, were tested for functional activity of the receptor binding domain in an ACE2 binding assay. For this, we used human HEK 293T +ACE2 +TMPRSS2 (hek+) cells that we were given to us by our sponsor VectorBuilder. This specific cell line has a mutation that renders an overexpression of the RBD receptor angiotensin I converting enzyme 2, short ACE2. As negative control we employed HEK 293T cells that don’t express ACE2 (hek-). For a specific protocol for the conduction of this activity assay utilizing human cell culture, please refer to our Experiments page.
For the activity assay seen in Figure 7, one well of HEK-cells was incubated with L1_sAP_RBD_Strep8His and another well with weird_plex,for both of which the cell culture supernatant was concentrated through ammonium sulfate precipitation. One further well of HEK-cells were incubated with the purified L1_sAP_RBD_GST. HEK- -cells were incubated as negative controls.
-
 Figure 7 | Immunoblot | stained against ACE2 and RBD.
Figure 7 | Immunoblot | stained against ACE2 and RBD.
10% SDS-gel | 20 µL loaded per sample
A | stained with ms α-ACE2 | 1:1000
B | stained with ms α-RBD | 1:2000
1 | HEK-- cells incubated with 200 µL RBD_TEV_GST | purified
2 | HEK+-cells incubated with 1.5 mL RBD_Strep8His | concentrated through ammonium sulfate precipitation
3 | HEK+-cells incubated with 1.5 mL weird_plex | to rule out cross reaction with Leishmania supernatant
4 | HEK+- cells incubated with 200 µL RBD_TEV_GST | purified
5 | HEK--cells incubated with 1.5 mL RBD_Strep8His | concentrated through ammonium sulfate precipitation
6 | HEK+-cells only
7 | HEK-cells only
L | Thermo Scientific PageRuler Protein Ladder

For the fusion protein RBD_Strep8His on HEK+ cells, we can see a band at the expected height (27 kDa), which is a first indication of functionality of our protein RBD. Furthermore, the band is not present in the negative control (Hek--RBD-Strep8His) where we incubated the HEK--cells with the same sample. This proves that the band is not a cross reaction of the antibody with any other proteins of the HEK cells or Leishmania supernatant.
The purified RBD_GST shows no such band. This could be because the RBD got lost during purification, or because the GST tag, which is as big as the RBD itself, structurally interferes with ACE2 binding. The HEK-cells that were incubated with weird_plex show, as expected, no RBD.
The MocloMania collection
This basic part is part of the MocloMania collection, the very first collection of genetic parts specifically designed and optimized for Modular Cloning assembly and recombinant protein expression in the protozoan parasite Leishmania tarentolae.
Are you trying to express complexly glycosylated proteins? Large antibody side chains? Human proteins that require accurate post-translational modification? Then Leishmania might be just the right organism for you! Leishmania tarentolae’s glycosylation patterns resemble those of human cells more closely than any other microbial expression host, while still delivering all the benefits of microbial production systems like easy transfection and cultivation.[5] So instead of relying on mammalian cell lines, try considering Leishmania as your new expression host of choice!
Our MocloMania collection will allow you to easily modify your protein of choice and make it suitable for downstream detection and purification procedures - all thanks to the help of Modular Cloning. This cloning system was first established by Weber et al. in 2011 and relies on the ability of type IIS restriction enzymes to cut DNA outside of their recognition sequence, hereby generating four nucleotide overhangs.[6] Every basic part in our collection is equipped with a specified set of overhangs that assign it to its designated position within the reading frame. These so-called cloning positions are labelled B2-B5 from upstream to downstream. By filling all positions with the basic parts of your choice, you can easily generate variable genetic constructs that code for the fusion protein of your desire.
We furthermore provide a specifically domesticated Leishmania expression vector, named weird_plex, which will package your fusion construct into a functional transcriptional unit that is optimized for high expression in Leishmania.
The best part? Because of the type IIS restriction properties and the specifity of the generated overhangs, restriction and ligation of your construct can all happen simultaneously in a simple one-step, one-pot reaction. This will safe you a lot of time and frustration in your cloning endeavours!
Do we have your attention? In the table below you can find some basic information on how our cloning system, along with most other MoClo systems, is set up. Please feel free to check out our wiki to find more information on Leishmania and Modular Cloning as well as to understand how this basic part integrates into our part collection. See you there!
| Level | What does this level contain? | antibiotic resistance | Enzyme used for ligation |
| L0 | The foundation to every MoClo construct which are basic genetic units, such as coding sequences, promoters, terminators | spectinomycin | BbsI |
| L1 | Several L0 parts assembled into a functional transcriptional unit, e.g. consisting of promoter, coding region and terminator | ampicillin | BsaI |
| L2 | Multiple transcriptional units added into one multi-gene construct, e.g. a protein of interest fused to a selection marker | kanamycin | BbsI |
Sequence and Features
IMPORTANT
This basic part has been altered in its sequence in order to make it compatible with the iGEM RCF 1000 cloning standard.
In its original form, this part contains an endogenous SapI restriction site. This is because it was specifically designed to be utilized within the Modular Cloning system as established by Weber et al. in 2011, relying on BsaI and BbsI as the executive type IIS restriction enzymes.[7] This MoClo standard is a very frequently used type IIS standard, especially when it comes to plant-based expression systems.[8][9]
Because of its intended use within the MoClo system, this basic part is domesticated against BsaI and BbsI, but not against SapI. Upon registration of this part in the iGEM Registry, the endogenous SapI restriction site was disrupted via in silico introduction of a silent mutation. We decided to do this in order to increase the chance of the judges actually taking a look at this part page and not having the part overlooked due to incompatibility.
Since the part’s amino acid sequence remains unchanged, users interested in utilizing this part for type IIS loop assembly can rely on the sequence as listed on this page. For users interested in applying this basic part to their MoClo based cloning approaches, in silico reversion of the mutation might be beneficial, since SapI proved to be a valuable enzyme for confirming part insertion via restriction digest.
We highly recommend that researchers and iGEM teams utilize this basic part within the MocloMania cloning system, since it is codon optimized towards Leishmania tarentolae and may otherwise negatively influence expression yield in other organisms.
MUTATION | bp 634 - 636 | AGC → TCG
- 10INCOMPATIBLE WITH RFC[10]Illegal PstI site found at 520
- 12INCOMPATIBLE WITH RFC[12]Illegal PstI site found at 520
- 21INCOMPATIBLE WITH RFC[21]Illegal BglII site found at 458
- 23INCOMPATIBLE WITH RFC[23]Illegal PstI site found at 520
- 25INCOMPATIBLE WITH RFC[25]Illegal PstI site found at 520
Illegal NgoMIV site found at 606 - 1000COMPATIBLE WITH RFC[1000]
Reference Literature
- ↑ Aronson JK (25 March 2020). "Coronaviruses – a general introduction". Centre for Evidence-Based Medicine, Nuffield Department of Primary Care Health Sciences, University of Oxford.
- ↑ F. Li, Structure, function, and evolution of coronavirus spike proteins. Annu. Rev. Virol. 3, 237–261
- ↑ Shang, J., Ye, G., Shi, K. et al. Structural basis of receptor recognition by SARS-CoV-2. Nature 581, 221–224 (2020). https://doi.org/10.1038/s41586-020-2179-y
- ↑ M. Hoffmann et al., SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280.e8 (2020).
- ↑ Langer T, Corvey C, Kroll K, Boscheinen O, Wendrich T, Dittrich W. Expression and purification of the extracellular domains of human glycoprotein VI (GPVI) and the receptor for advanced glycation end products (RAGE) from Rattus norvegicus in Leishmania tarentolae. Prep Biochem Biotechnol. 2017 Nov 26;47(10):1008-1015. doi: 10.1080/10826068.2017.1365252. Epub 2017 Aug 31. PMID: 28857681.
- ↑ Weber E, Engler C, Gruetzner R, Werner S, Marillonnet S (2011) A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLoS ONE 6(2): e16765. https://doi.org/10.1371/journal.pone.0016765
- ↑ Weber E, Engler C, Gruetzner R, Werner S, Marillonnet S (2011) A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLoS ONE 6(2): e16765. https://doi.org/10.1371/journal.pone.0016765
- ↑ Patron, N.J., Orzaez, D., Marillonnet, S. et al., (2015), Standards for plant synthetic biology: a common syntax for exchange of DNA parts. New Phytol, 208: 13-19. https://doi.org/10.1111/nph.13532
- ↑ https://www.addgene.org/cloning/moclo/, last visited 10/16/21, 19:00 ECT
| biology | SARS-CoV2 |
| function | receptor binding to mediate viral cell entry |
| ligands | Angiotensin Converting Enzyme 2 (ACE2) |