
Part:BBa_K3781016
mCerulean, MocloMania B5
This part codes for the cyan fluorescent protein mCerulean.[1] The protein was originally derived from the green fluorescent protein GFP which was first isolated from the bioluminescent jellyfish Aequorea victoria in 1962.[2] It can be fused to a protein of interest, making it accessible to easy and non-invasive screening opportunities that involve fluorescence monitoring, such as fluorescence spectroscopy or microscopy.[3] As a B5 part, this part is meant to occupy the most downstream position in the MocloMania cloning frame and is thus equipped with an additional stop codon.
size 26.8 kDa
function fluorescent tag
excitation wavelength 435 nm
emission wavelength 477 nm
cloning position B5
plasmid backbone pAGM1301
Data
We were able to successfully clone this basic part into its respective L0 plasmid backbone and to confirm the integrity of the L0 construct via restriction digest and gel electrophoresis, see Figure 1. Furthermore, we were able to include it into a L1 construct, proving its correct adaptation towards MoClo assembly, see Figure 2.
-
 Figure 1 | Test digest of L0 B5 parts using BsaI
Figure 1 | Test digest of L0 B5 parts using BsaI
1 | pAGM1301 | 2247 + 598 bp
2 | L0_mCerulean_B5 | 2247 + 721 bp
3 | L0_mVenus_B5 | 2247 + 721 bp
4 | L0_GST_B5 | 2247 + 688 bp
L | Thermofischer GeneRuler Plus Ladder [bp] -
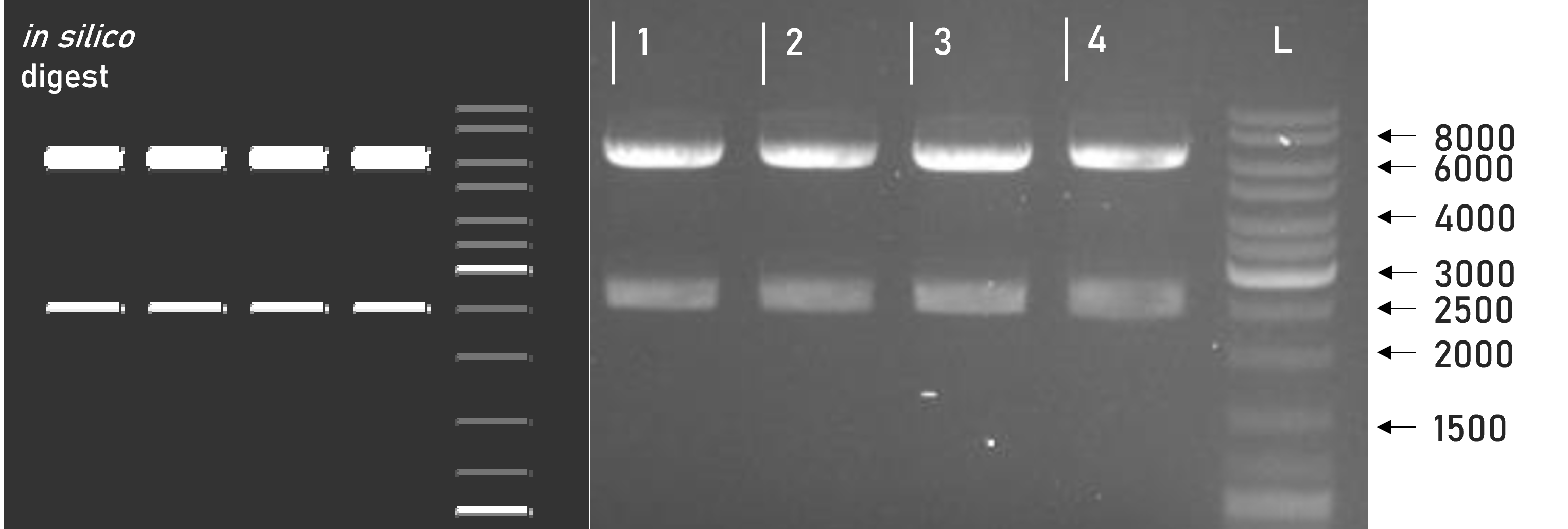 Figure 2 | Test digest of L1 constructs using SacI
Figure 2 | Test digest of L1 constructs using SacI
1 | L1_3xHA_RBD_mVenus | 6098 + 2515 bp
2 | L1_3xHA_RBD_mCerulean | 6098 + 2515 bp
3 | L1_sAP_RBD_mVenus | 6071 + 2515 bp
4 | L1_sAP_RBD_mCerulean | 6071 + 2515 bp
L | Thermofischer GeneRuler Plus Ladder [bp]


Several different L1 constructs assembled with L0_mCerulean_B5 have been successfully transfected into Leishmania, resulting in recombinant protein expression that could be observed after immunostaining on western blot.
-
 Figure 3 | Immunoblot of L1 transfected Leishmania | stained against RBD
Figure 3 | Immunoblot of L1 transfected Leishmania | stained against RBD
1 | L1_sAP_RBD_mCerulean | 51.9 kDa
2 | L1_sAP_RBD_mCerulean_Strep8His | 54.3 kDa
3 | L1_sAP_RBD_mCerulean_GST | 78.6 kDa
4 | L1_3xHA_RBD_mCerulean | 55.7 kDa
n.c. | negative control | Leishmania culture
transfected with empty L1 expression vector p.c. | RBD-GFP | 54 kDa
L | Thermofischer PageRuler Protein Ladder [kDa]
1. AB | ms anti-RBD
2. AB | rb anti-ms HRP

Looking at Figure 3, we see that both constructs that employ L0_mCerulean_B5, namely 1 | L1_sAP_RBD_mCerulean and 4 | L1_3xHA_RBD_mCerulean, show definite protein bands when stained against the SARS-CoV-2 receptor binding domain. As construct 1 contains the sAP secretion tag, we can see that protein expression is detected both in the cell lysate P+ as well as in the culture supernatant S+. Judging from the signal intensity, levels of secreted protein exceed those detected within the lysate. Here, part of the recombinant protein might still have been intracellularly processed or got stuck along the secretory pathway, e.g. during membrane passage. On the other hand, construct 4 is not equipped with a secretion tag and, consequently, its expression can only be detected in the cell lysate.
What is remarkable is that according to in silico calculations, we expect both constructs to weigh about 50 kDa. Taking into consideration small deficiencies in gel density due to high percentage of acrylamide, both constructs' upper lanes can be attributed to that size. However, the secretion of construct 1 into the culture medium seems to initiate a protein cleavage process, resulting in a lower band that runs no higher than 28 kDa. This lower band can be seen in all secreted constructs and exclusively in secreted constructs. Its visualisation with the help of anti-RBD antibodies as well as its size suggest a possible post-translational cleavage process in the cell culture medium separating the receptor binding domain from its fused protein tags. This issue could be tackled by introducing protease inhibitors into the culture mix after induction of recombinant protein expression.
-
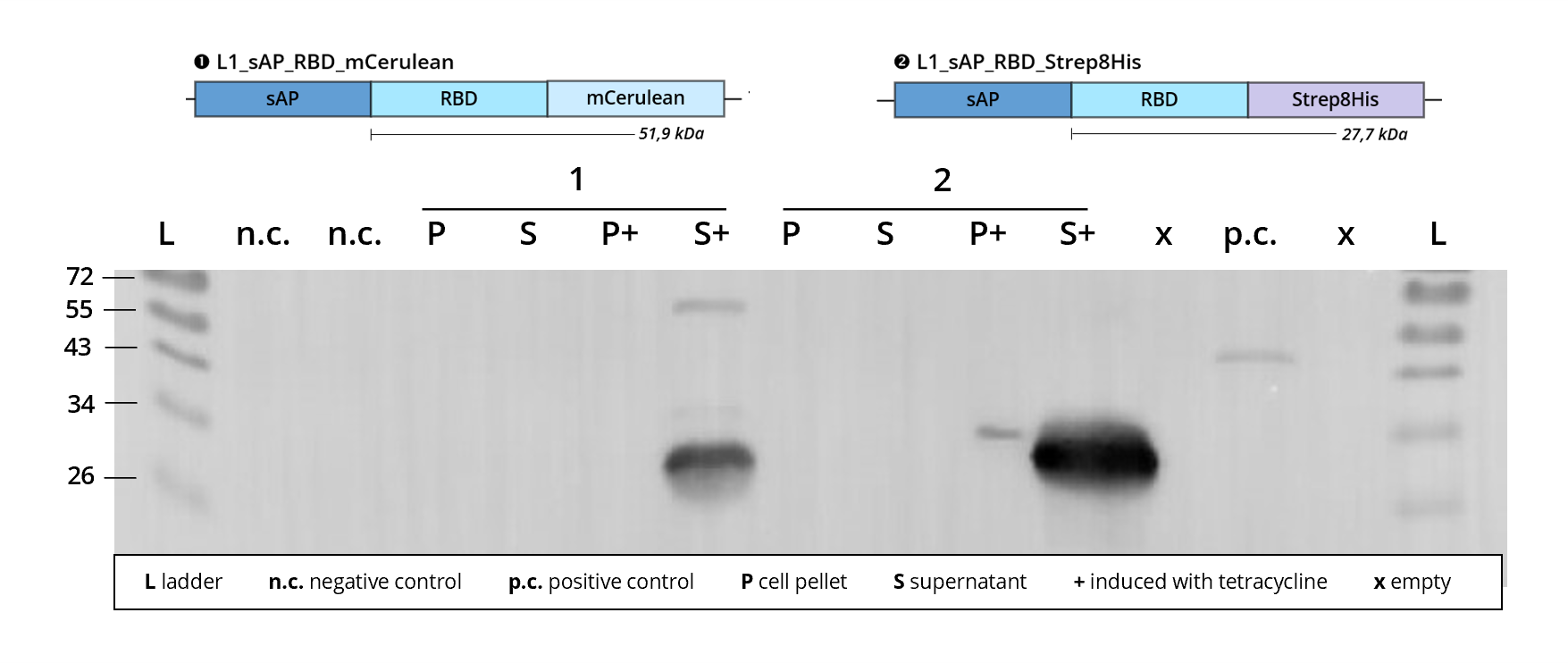 Figure 4 | Immunoblot of L1 transfected Leishmania | stained against RBD
Figure 4 | Immunoblot of L1 transfected Leishmania | stained against RBD
1 | L1_sAP_RBD_mCerulean | 51.9 kDa
2 | L1_sAP_RBD_Strep8His | 27.7 kDa
n.c. | negative control | untransfected Leishmania culture
p.c. | RBD-GFP | 54 kDa
L | Thermofischer PageRuler Protein Ladder [kDa]
1. AB | ms anti-RBD
2. AB | rb anti-ms HRP

The same observation can be made when looking at Figure 4, where again the upper band of 1 | L1_sAP_RBD_Strep8His runs very sharply and within the speculated 50 kDa range and the lower band gives a much more smeared band at around 28 kDa. For this transfected cell culture, protein expression is exclusively detected in the cell culture supernatant. Beyond that, the blot in Figure 4 as well as the first sample in Figure 3 visualize the functionality of the tetracycline induced transcription activation, a feature of the MocloMania L1 expression vector weird_plex. Whereas before tetracycline addition, S and P, the cultures display almost no detectable protein expression, the induction process, S+ and P+, results in strong and defined protein bands.
So far, none of the L1 constructs employing a C-terminal mCerulean have been successfully purified. Furthermore, fluorescence screenings of cells producing recombinant mCerulean have not yet been conducted. Thus, at this point in time, functionality of the mCerulean fusion tag cannot be definitively verified. We are working hard on optimizing expression and purification yields in order to finalize part validation and implementation towards their respective functionality.
The MocloMania collection
This basic part is part of the MocloMania collection, the very first collection of genetic parts specifically designed and optimized for Modular Cloning assembly and recombinant protein expression in the protozoan parasite Leishmania tarentolae.
Are you trying to express complexly glycosylated proteins? Large antibody side chains? Human proteins that require accurate post-translational modification? Then Leishmania might be just the right organism for you! Leishmania tarentolae’s glycosylation patterns resemble those of human cells more closely than any other microbial expression host, while still delivering all the benefits of microbial production systems like easy transfection and cultivation.[4] So instead of relying on mammalian cell lines, try considering Leishmania as your new expression host of choice!
Our MocloMania collection will allow you to easily modify your protein of choice and make it suitable for downstream detection and purification procedures - all thanks to the help of Modular Cloning. This cloning system was first established by Weber et al. in 2011 and relies on the ability of type IIS restriction enzymes to cut DNA outside of their recognition sequence, hereby generating four nucleotide overhangs.[5] Every basic part in our collection is equipped with a specified set of overhangs that assign it to its designated position within the reading frame. These so-called cloning positions are labelled B2-B5 from upstream to downstream. By filling all positions with the basic parts of your choice, you can easily generate variable genetic constructs that code for the fusion protein of your desire.
We furthermore provide a specifically domesticated Leishmania expression vector, named weird_plex, which will package your fusion construct into a functional transcriptional unit that is optimized for high expression in Leishmania.
The best part? Because of the type IIS restriction properties and the specifity of the generated overhangs, restriction and ligation of your construct can all happen simultaneously in a simple one-step, one-pot reaction. This will safe you a lot of time and frustration in your cloning endeavours!
Do we have your attention? In the table below you can find some basic information on how our cloning system, along with most other MoClo systems, is set up. Please feel free to check out our wiki to find more information on Leishmania and Modular Cloning as well as to understand how this basic part integrates into our part collection. See you there!
| Level | What does this level contain? | antibiotic resistance | Enzyme used for ligation |
| L0 | The foundation to every MoClo construct which are basic genetic units, such as coding sequences, promoters, terminators | spectinomycin | BbsI |
| L1 | Several L0 parts assembled into a functional transcriptional unit, e.g. consisting of promoter, coding region and terminator | ampicillin | BsaI |
| L2 | Multiple transcriptional units added into one multi-gene construct, e.g. a protein of interest fused to a resistance cassette | kanamycin | BbsI |
Sequence and Features
- 10COMPATIBLE WITH RFC[10]
- 12COMPATIBLE WITH RFC[12]
- 21COMPATIBLE WITH RFC[21]
- 23COMPATIBLE WITH RFC[23]
- 25INCOMPATIBLE WITH RFC[25]Illegal NgoMIV site found at 679
- 1000COMPATIBLE WITH RFC[1000]
Reference Literature
- ↑ R Heim, D C Prasher, R Y Tsien, "Wavelength mutations and posttranslational autoxidation of green fluorescent protein", Proceedings of the National Academy of Sciences Dec 1994, 91 (26) 12501-12504; DOI: 10.1073/pnas.91.26.12501
- ↑ Shimomura, O., Johnson, F.H. and Saiga, Y. (1962) Extraction, Purification, and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan, Aequorea. Journal of Cellular and Comparative Physiology, 59, 223-239. http://dx.doi.org/10.1002/jcp.1030590302
- ↑ M.A. Rizzo, D.W. Piston High-contrast imaging of fluorescent protein FRET by fluorescence polarization microscopy Biophys. J., 88 (2005), pp. L14-L16
- ↑ Langer T, Corvey C, Kroll K, Boscheinen O, Wendrich T, Dittrich W. Expression and purification of the extracellular domains of human glycoprotein VI (GPVI) and the receptor for advanced glycation end products (RAGE) from Rattus norvegicus in Leishmania tarentolae. Prep Biochem Biotechnol. 2017 Nov 26;47(10):1008-1015. doi: 10.1080/10826068.2017.1365252. Epub 2017 Aug 31. PMID: 28857681.
- ↑ Weber E, Engler C, Gruetzner R, Werner S, Marillonnet S (2011) A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLoS ONE 6(2): e16765. https://doi.org/10.1371/journal.pone.0016765
//function/reporter/fluorescence
| biology | cyan fluorescent protein |
| emission | 435 nm |
| excitation | 477 nm |