
Difference between revisions of "Part:BBa K1586000"
| (9 intermediate revisions by the same user not shown) | |||
| Line 18: | Line 18: | ||
<br /> | <br /> | ||
In order to characterise that this part works as expected, fluorescence intensity was measured for the expression of K1586000 in a cell free system in the presence of different amounts of trigger RNA, where the plasmid encoding for the toehold was kept constant at 0.5 pmols. Below is a graph showing a positive correlation between amount of trigger RNA (in log10(nanograms) with 0ng normalised to 0) and GFP fluorescence intensity. | In order to characterise that this part works as expected, fluorescence intensity was measured for the expression of K1586000 in a cell free system in the presence of different amounts of trigger RNA, where the plasmid encoding for the toehold was kept constant at 0.5 pmols. Below is a graph showing a positive correlation between amount of trigger RNA (in log10(nanograms) with 0ng normalised to 0) and GFP fluorescence intensity. | ||
| + | <br /> | ||
https://static.igem.org/mediawiki/2015/5/5a/Exeter_GreenJ_trigger_conc_graph.png | https://static.igem.org/mediawiki/2015/5/5a/Exeter_GreenJ_trigger_conc_graph.png | ||
| − | + | <br /> | |
| + | <br /> | ||
| + | <br /> | ||
| + | <br /> | ||
| + | <br /> | ||
| + | <br /> | ||
| + | It should be noted that this graph was generated from a single repeat due to limitations in testing time and the cost of cell-free reactions. While this graph does show a general trend upwards, the lack of repeats and therefore error bars means that these results should be treated with caution. However, additional data (shown below) does support the conclusion which we have drawn here. | ||
<html> | <html> | ||
| − | <b>"In silico | + | |
| + | <b>Specificity:</b> | ||
| + | </br> | ||
| + | In addition to this, GreenFET1J was expressed in the presence of a random incorrect trigger RNA sequence. As can be seen from the graph below, the GFP production from GreenFET1J in presence of correct trigger RNA is much higher when compared to GFP production in the presence of incorrect trigger RNA. This shows that the toehold is specific towards the correct trigger RNA. | ||
| + | |||
| + | <img src="https://static.igem.org/mediawiki/parts/7/7b/Exeter_specificity_of_toehold.png" width="700px"> | ||
| + | </br> | ||
| + | </br> | ||
| + | </br> | ||
| + | </br> | ||
| + | |||
| + | As with the data above, these results should be treated with caution as the cell-free positive control ran with this experiment was negative. Despite this, the data above is included in the characterisation of this part as the other reactions behaved as expected, and they are also in line with the data posted above, suggesting that the conclusions drawn are correct. | ||
| + | |||
| + | |||
| + | <b><em>In silico</em>:</b> | ||
<br /> | <br /> | ||
| − | As well as generating experimental data in the lab, | + | As well as generating experimental data in the lab, <em>in silico</em> data was also generated through the software package <a href="http://www.nupack.org/">NUPACK</a> using free energies. For a full explanation of how this works, please visit http://2015.igem.org/Team:Exeter/Modeling.</br> |
</br> | </br> | ||
The equilibrium concentrations of the different components in our system were determined, along with free energy data for the different structures; the unbound trigger, the unbound toehold and the complex of the two. | The equilibrium concentrations of the different components in our system were determined, along with free energy data for the different structures; the unbound trigger, the unbound toehold and the complex of the two. | ||
| − | <img | + | <img src="https://static.igem.org/mediawiki/2015/9/93/FreeEnergiesNupackGraph.png" width="700px"> |
| − | + | </br> | |
| + | </br> | ||
| + | </br> | ||
| + | </br> | ||
| + | </br> | ||
After obtaining this, the data was converted to an approximate normal distribution and after normalising these within the range of values, we could use the distribution as a probability distribution. Shown below are these obtained probability distributions based on the free energy data. | After obtaining this, the data was converted to an approximate normal distribution and after normalising these within the range of values, we could use the distribution as a probability distribution. Shown below are these obtained probability distributions based on the free energy data. | ||
| Line 37: | Line 62: | ||
<img src="https://static.igem.org/mediawiki/2015/d/df/NormalisedBindingProbabilityGraph.png" width="700px" | <img src="https://static.igem.org/mediawiki/2015/d/df/NormalisedBindingProbabilityGraph.png" width="700px" | ||
| − | + | </br> | |
| + | </br> | ||
<img src="https://static.igem.org/mediawiki/2015/a/a0/NormalisedSplittingProbability.png" width="700px"> | <img src="https://static.igem.org/mediawiki/2015/a/a0/NormalisedSplittingProbability.png" width="700px"> | ||
Latest revision as of 00:05, 27 September 2015
Synthetic toehold riboswitch - J23100
Usage and Biology
A toehold switch is a type of RNA molecule known as a riboregulator/riboswitch. It is able to detect the presence of a specific ssRNA molecule (termed the 'trigger RNA') which has a sequence complementary to its switch region through base pairing. If the correct RNA molecule is detected, the protein coding region of the toehold is expressed.
A toehold switch is unique in comparison to other types of riboswitches as it is completely synthetic, and therefore easier to engineer and standardise. The fact that the toehold switch can be modified means that the switch region can be easily changed to detect any given trigger RNA molecule, and the protein coding region can be swapped for any desired reporter protein most suitable for its application.
As shown by Exeter iGEM 2015, toehold switches can be used to detect specific RNA molecules in a cell-free system. In addition to this, the plasmid DNA encoding the toehold switch can be transformed into cells in order to ascertain whether a gene is being expressed (through detection of its mRNA). The applications of this technology can range from a research tool (e.g. detection of secreted RNA in cell supernatant, detection of gene expression, etc.), through to more commercial/medical applications such as diagnostic testing.
The Part
This part encodes for a synthetic toehold switch which has been modified to be under the control of a standard BBa_J23100 constitutive promoter. Originally, this toehold also contained an illegal PstI site at position 844, however this was removed to allow standardisation into a biobrick.
Characterisation
Experimental:
In order to characterise that this part works as expected, fluorescence intensity was measured for the expression of K1586000 in a cell free system in the presence of different amounts of trigger RNA, where the plasmid encoding for the toehold was kept constant at 0.5 pmols. Below is a graph showing a positive correlation between amount of trigger RNA (in log10(nanograms) with 0ng normalised to 0) and GFP fluorescence intensity.

It should be noted that this graph was generated from a single repeat due to limitations in testing time and the cost of cell-free reactions. While this graph does show a general trend upwards, the lack of repeats and therefore error bars means that these results should be treated with caution. However, additional data (shown below) does support the conclusion which we have drawn here.
Specificity:
In addition to this, GreenFET1J was expressed in the presence of a random incorrect trigger RNA sequence. As can be seen from the graph below, the GFP production from GreenFET1J in presence of correct trigger RNA is much higher when compared to GFP production in the presence of incorrect trigger RNA. This shows that the toehold is specific towards the correct trigger RNA.
 As with the data above, these results should be treated with caution as the cell-free positive control ran with this experiment was negative. Despite this, the data above is included in the characterisation of this part as the other reactions behaved as expected, and they are also in line with the data posted above, suggesting that the conclusions drawn are correct.
In silico:
As with the data above, these results should be treated with caution as the cell-free positive control ran with this experiment was negative. Despite this, the data above is included in the characterisation of this part as the other reactions behaved as expected, and they are also in line with the data posted above, suggesting that the conclusions drawn are correct.
In silico:
As well as generating experimental data in the lab, in silico data was also generated through the software package NUPACK using free energies. For a full explanation of how this works, please visit http://2015.igem.org/Team:Exeter/Modeling.
The equilibrium concentrations of the different components in our system were determined, along with free energy data for the different structures; the unbound trigger, the unbound toehold and the complex of the two.
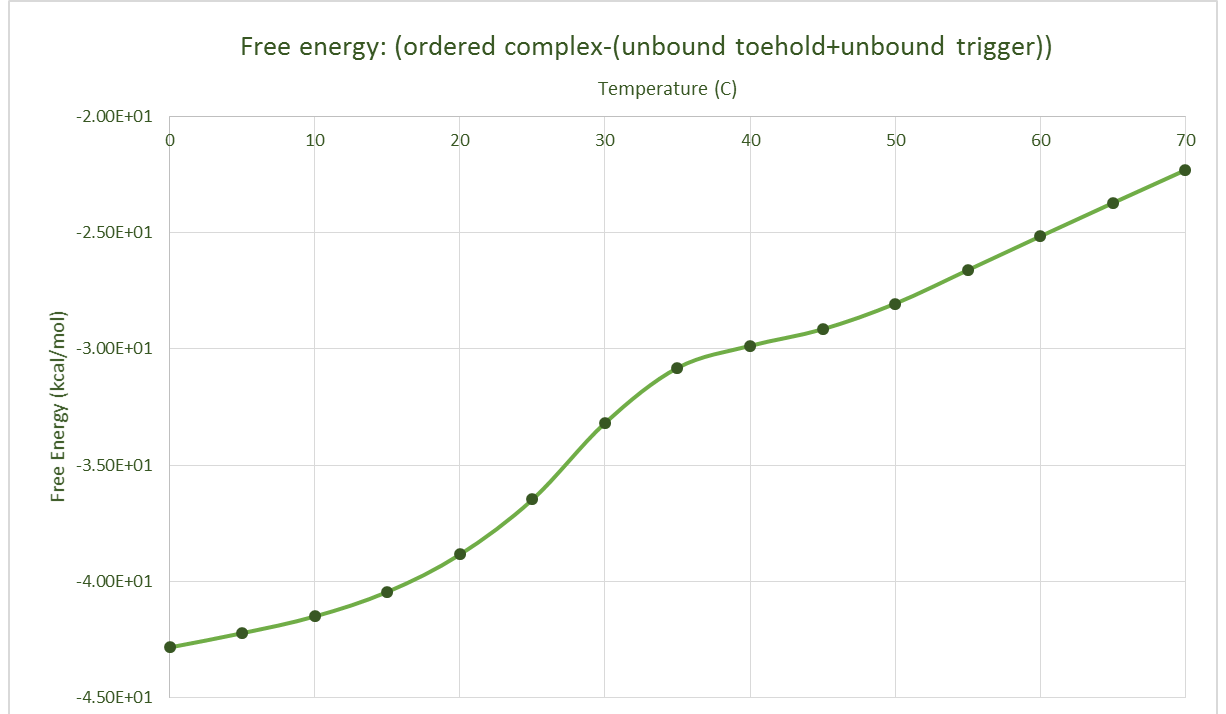 After obtaining this, the data was converted to an approximate normal distribution and after normalising these within the range of values, we could use the distribution as a probability distribution. Shown below are these obtained probability distributions based on the free energy data.
After obtaining this, the data was converted to an approximate normal distribution and after normalising these within the range of values, we could use the distribution as a probability distribution. Shown below are these obtained probability distributions based on the free energy data.
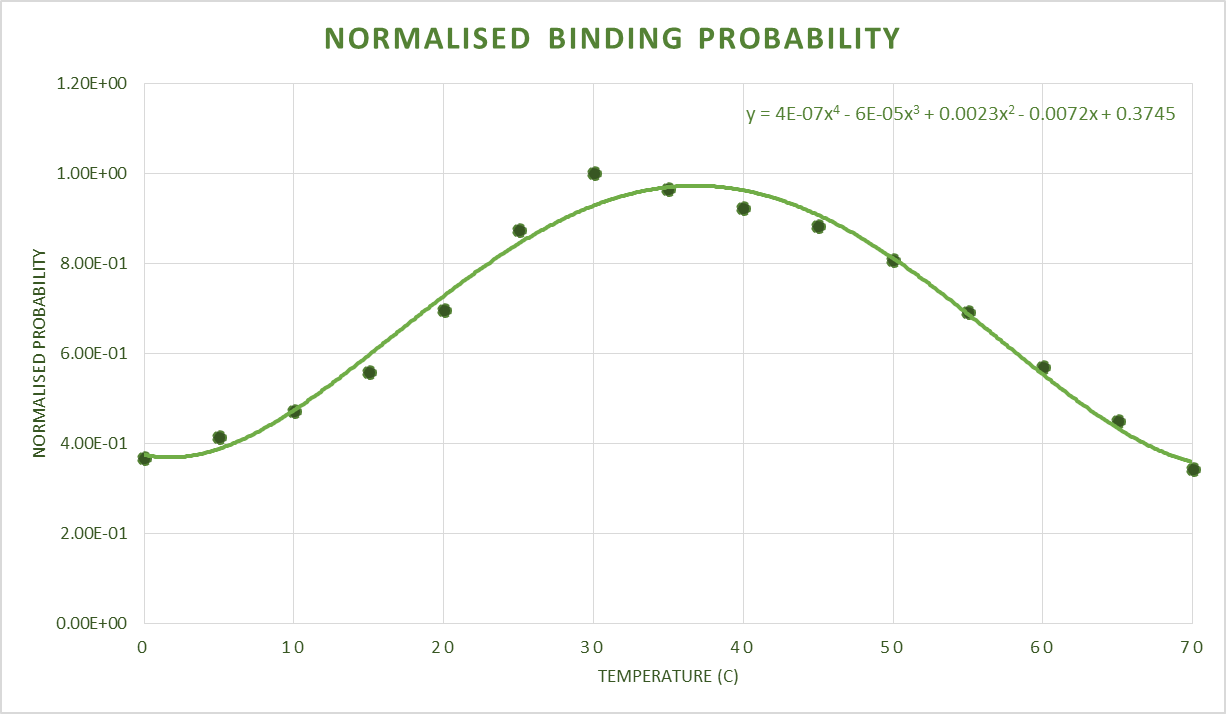
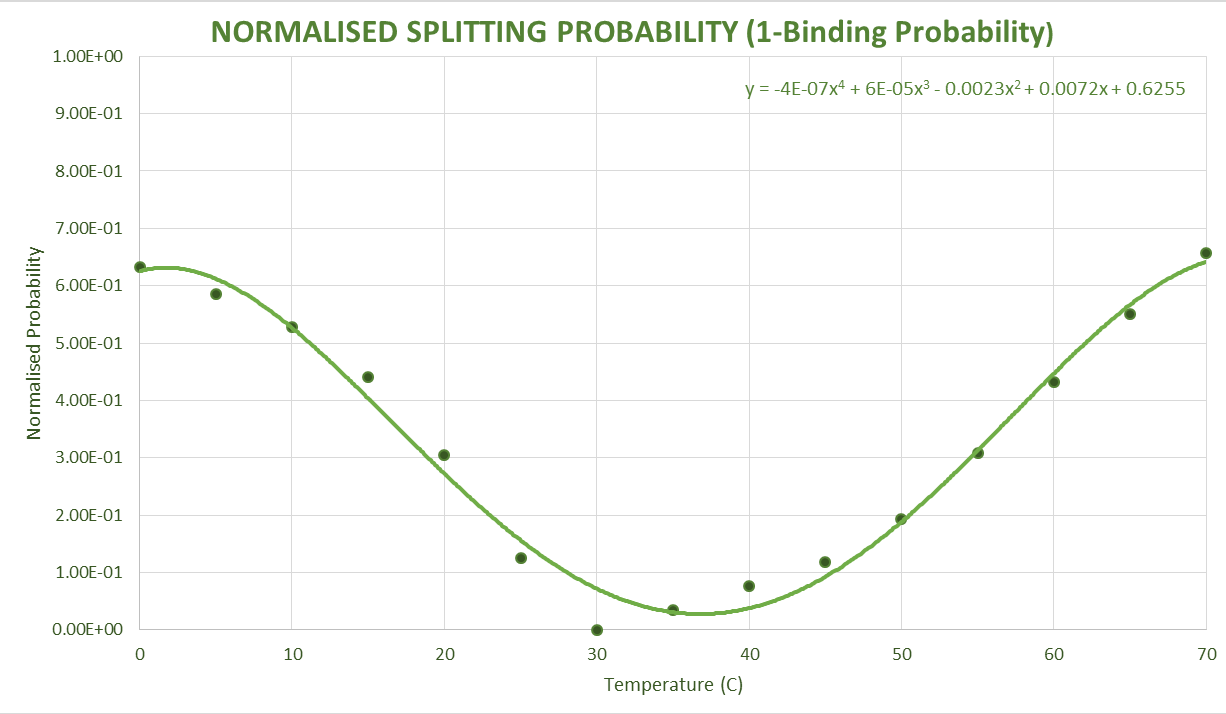
Sequence and Features
- 10COMPATIBLE WITH RFC[10]
- 12INCOMPATIBLE WITH RFC[12]Illegal NheI site found at 7
Illegal NheI site found at 30 - 21COMPATIBLE WITH RFC[21]
- 23COMPATIBLE WITH RFC[23]
- 25COMPATIBLE WITH RFC[25]
- 1000INCOMPATIBLE WITH RFC[1000]Illegal BsaI.rc site found at 769