Designed by: Ina Schmitt Group: iGEM19_Bielefeld-CeBiTec (2019-10-15)
Kanamycin resistance protein Part 1
Sequence and Features
Assembly Compatibility:
10
COMPATIBLE WITH RFC[10]
12
COMPATIBLE WITH RFC[12]
21
COMPATIBLE WITH RFC[21]
23
COMPATIBLE WITH RFC[23]
25
COMPATIBLE WITH RFC[25]
1000
COMPATIBLE WITH RFC[1000]
Summary
Would you love working with a two-plasmid system using only one antibiotic resistance? We enable you to do this by splitting antibiotic resistance genes with intein-mediated protein trans-splicing. By implementing this innovative approach, the likelihood of spreading resistances is decreased as one plasmid does not encode the full protein. Therefore, a single plasmid is insufficient to confer an antibiotic resistance to bacteria in the environment, functioning as a biosafety system. To ensure the universal applicability within the iGEM community we selected resistances against commonly used antibiotics: Chloramphenicol, Kanamycin, Ampicillin and Hygromycin B. The optimal split point was selected based on our model, taking into consideration the 3D-structure and the amino acids relevant for the splicing process. By constructing and characterizing a collection of two-plasmid systems we demonstrated the feasibility of our split-antibiotic system to maintain two plasmids in a single cell. This intein-based approach is transferable to other resistance markers.
Another antibiotic used to prevent bacterial infections poses ampicillin. Similar to other antibiotics, it can be administrated via mouth and injection into a muscle. It is classified under the category of bacteriolytic antibiotics and can penetrate gram-positive and gram-negative bacteria. It is responsible for the inhibition of transpeptidase, that is responsible for the cell wall of the bacteria by inhibiting the final cell wall synthesis in binary fission (Ampicillin Monograph for Professionals, 2018)
Resistance Ampicillin
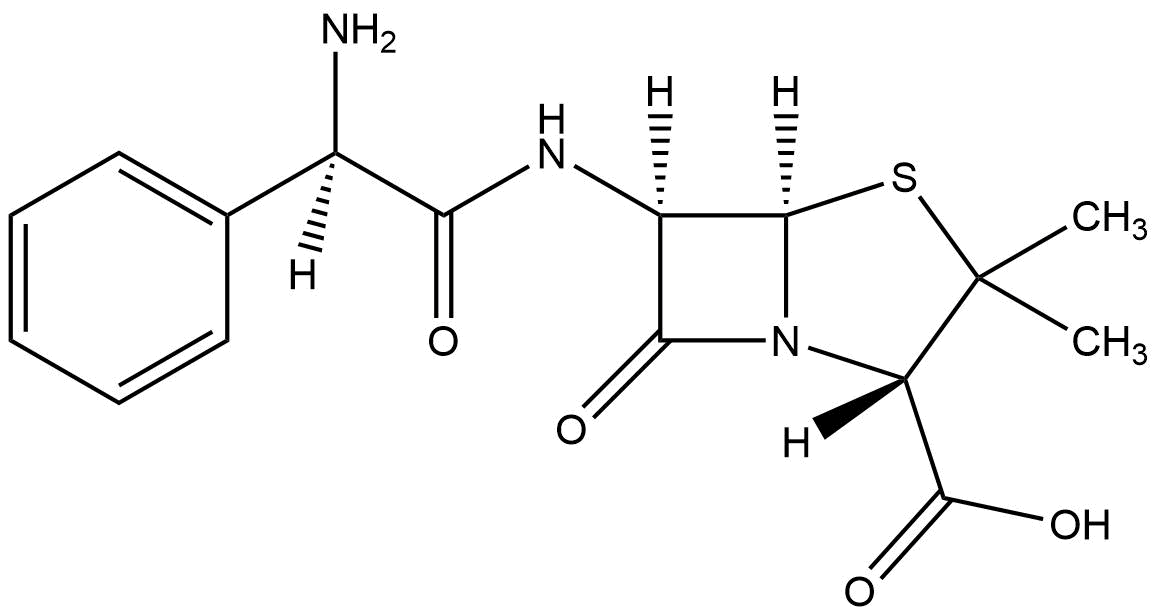
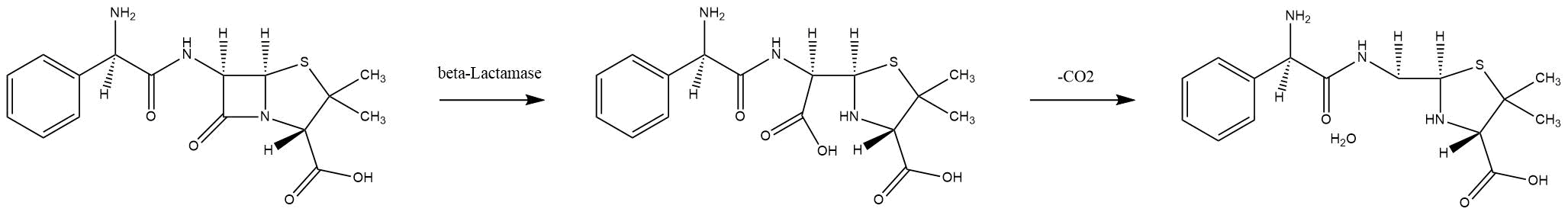
Beta-lactamases are enzymes that bacteria produce to defend themselves against β-lactam antibiotics. The most common lactamases are the class A enzymes, such as the clinically significant TEM-1 lactamase. (Waley, 1992). It provides antibiotic resistance by breaking the antibiotics structure. All these antibiotics all have a common element in their molecular structure: a four-atom ring known as a β-lactam. Through hydrolysis, the enzyme breaks the β-lactam ring open, deactivating the molecule's antibacterial properties.
Mechanism of TEM-1 lactamase hydrolysing ampicillin
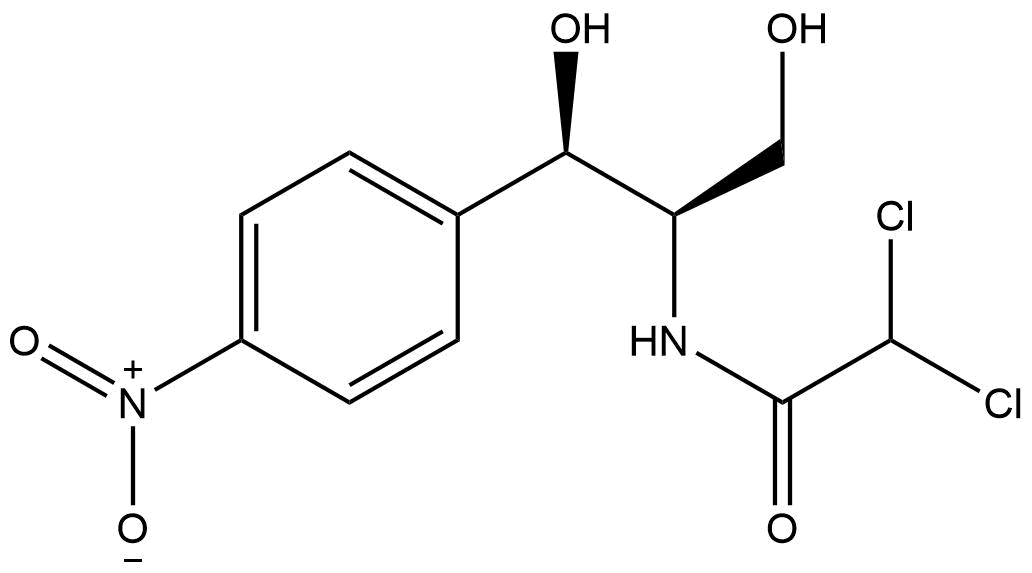
Chloramphenicol
Chemical structure of chloramphenicol
A commonly used antibiotic for bacterial infections is chloramphenicol (Chloramphenicol Monograph for Professionals, 2019). It was isolated from Streptomyces venezuelae in 1947 and was the first artificially produced antibiotic (Pongs O.,1979). Once it passes the cell membrane it has a bacteriolytic effect. It binds to specific residues of the 23S rRNA of the 50S ribosomal subunit, that hampers the substrate binding in the ribosome (Chloramphenicol - Infectious Diseases, 2019) (Wolfe, A. D., 1965). Chloramphenicol is often one of the ingredients in eye ointments for the treatment of conjunctivitis (Edwards 2009). In some other cases it can be administered by mouth or vein injection (Rosenfield, Logan, & Edwards, 2009). Furthermore, it is used to treat hazardous bacterial infections like plaque, cholera, MRSA or typhoid fever (Ingebrigtsen 2017).
Ribbon structure of the chloramphenicol acetyltransferase trimer chloramphenicol bound to the catalytic center. (From PDB: 3CLA)(Leslie, A.G.W. 1990)
Resistance Chloramphenicol
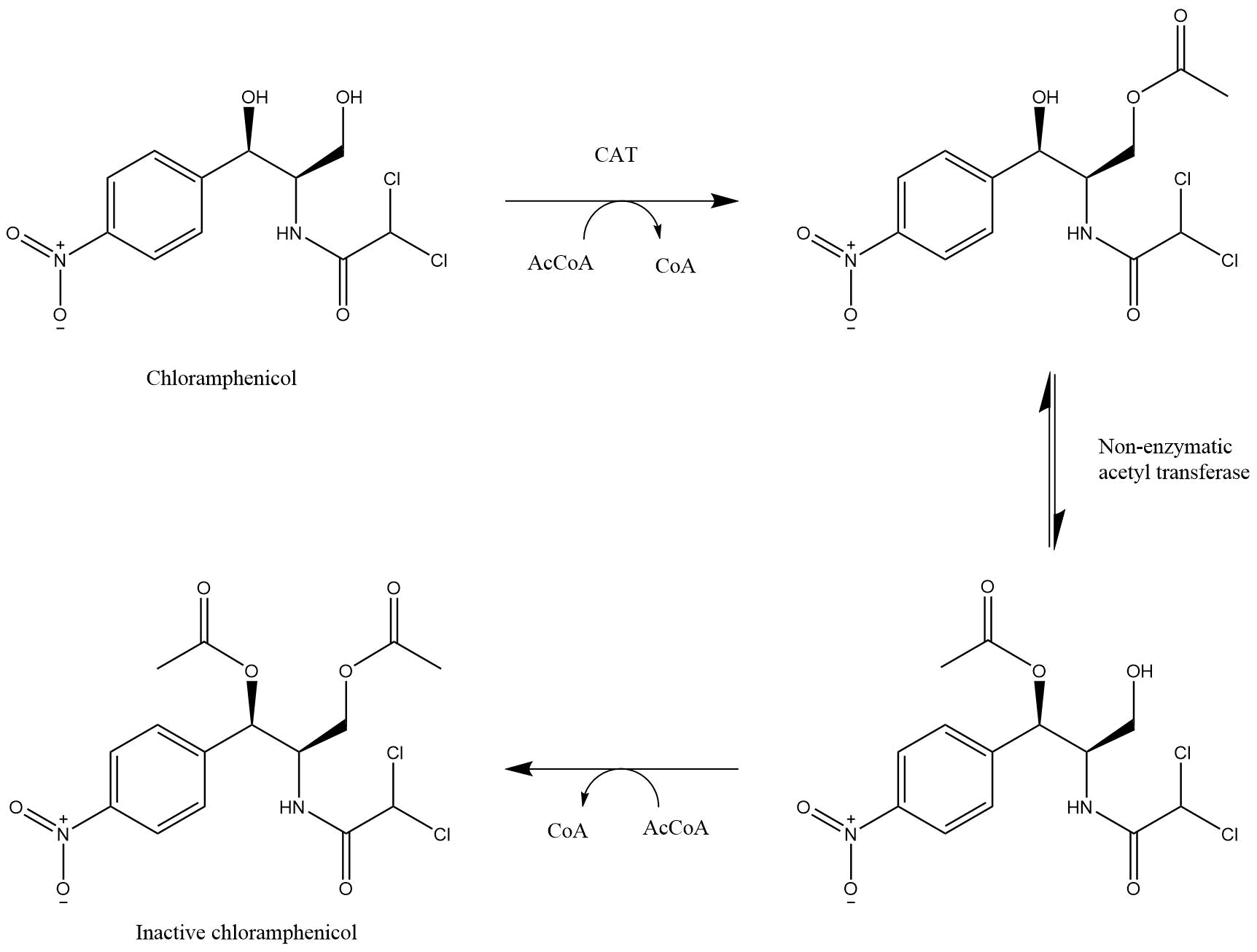
Chloramphenicol Acetyltransferase (CAT) is an enzyme originally identified in Escherichia coli that mediates resistance to chloramphenicol (Figure 3). (Shaw 1983). Three types of this enzyme are known and catalyse the same reaction, CATI, CATII and CATIII. The genomic analysis of the different CAT types shows that the boundaries between them are not completely clear, however type III is best characterised. Since the iGEM community uses commonly type I, as in part BBa_J31005 and in the standard backbone pSB1C3, we decided to do the same.
CAT is an enzyme originally identified in Escherichia coli that mediates resistance to chloramphenicol (Shaw 1983). It covalently attaches an acetyl group from acetyl-CoA to chloramphenicol, hence chloramphenicol is unable to bind to the 23S rRNA (Shaw 1991) (Figure 4)
Chemical structures of chloramphenicol and its acetylation by chloramphenicol acetyltransferase.
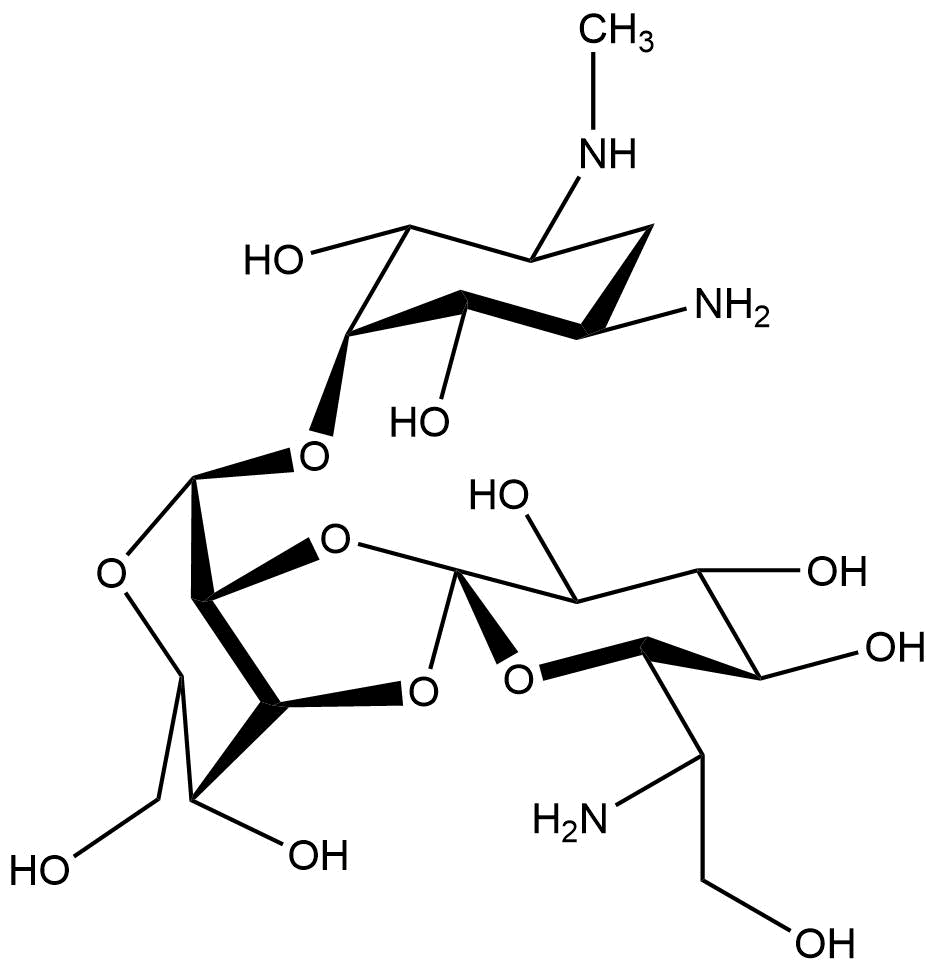
Hygromycin B
Chemical structure of hygromycin B
Hygromycin B is an aminoglycoside that can kill prokaryotic and eukaryotic cells. It was developed in 1950 and used for animals as an anti-worming agent. This antibiotic is produced by Streptomyces hygroscopicus and later in 1980 the resistance gene was discovered (Gritz & Davies, 1983; Kaster, Burgett, Rao, & Ingolia, 1983). The mechanism of hygromycin B is to inhibit the enzymic translocation of peptidyl-tRNA and thus blocking the cytoplasmic protein synthesis (González, Jiménez, Vázquez, Davies, & Schindler, 1978).
Resistance Hygromycin B
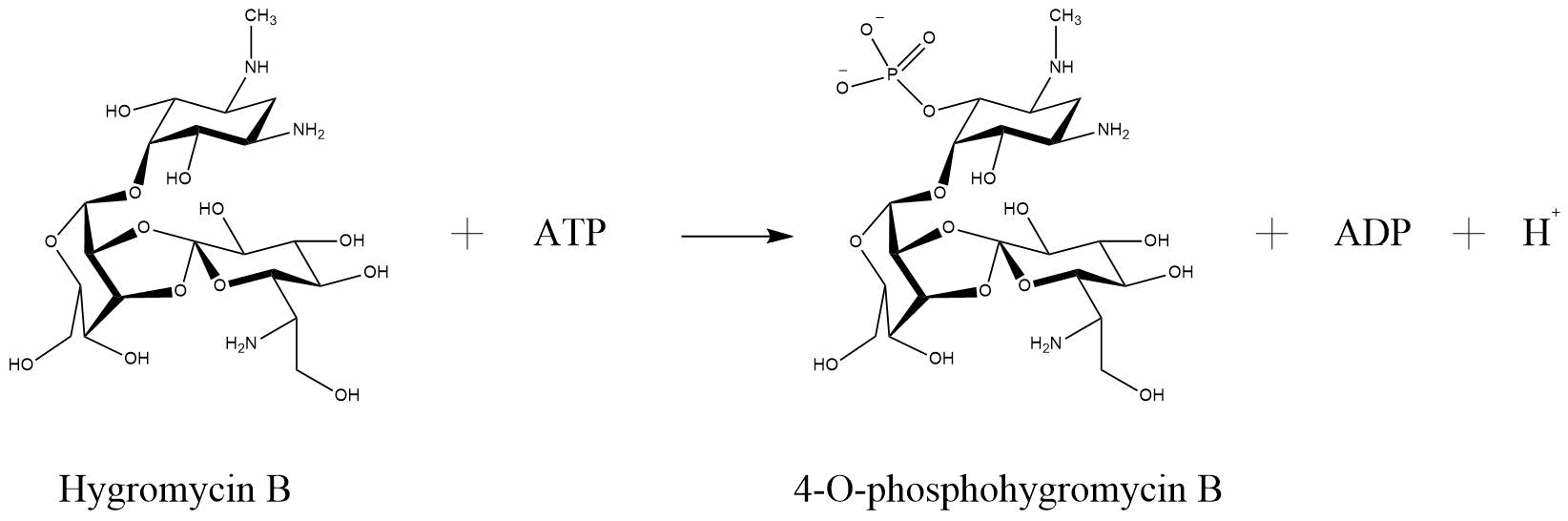
One of the enzymes that mediates resistance against hygromycin B (hygB) is called hygromycin-B 4-O-kinase and was found in E. coli (Hph—Hygromycin-B 4-O-kinase, Uniprot). It has two substrates, both ATP and Hygromycin B. It inactivates the antibiotic through phosphorilation, so that it has as products phospho-hygromycin B and ADP (Figure ). The same enzyme with the same characteristics, but from the organism Streptomyces fradiae named APH(4)-Ia, was tested for numerous aminoglycoside antibiotics, that perform possible substrates (Stogios et. al., 2011). The substrates that were tested included 4,6-disubstituted 2-deoxystreptamine-based, 4,5-disubstituted 2-deoxystreptamine-based and atypical aminoglycosides. The only antibiotic that was susceptible for phosphorylation by APH(4)-Ia was hygB. Such a specificity is not common for APHs in generall (Stogios et. al., 2011).
Reaction of hygromycin B become inactivated through phosphorylation via hygromycin-B 4-O-kinase (Hph—Hygromycin-B 4-O-kinase, Uniprot)
Kanamycin
Chemical structure of hygromycin B
Kanamycin is a glycoside that was first isolated in 1957 from the bacterium Streptomyces Kanamycericus (Sneader, 2005). It is used to treat bacterial infections and tuberculosis (‘Kantrex—FDA prescribing information, side effects and uses’, n.d.) and can be taken up by mouth, injection into a vein or into a muscle, but is ineffective against viral infections. The mechanism how kanamycin inhibit cell growth is by binding to the 30S subunit of the bacterial ribosome. This leads to incorrect alignment with the mRNA and therefore to the assembly of a false amino acid sequence. That leads to the assembly of on functional peptide chains (‘Kanamycin’, n.d.).
Resistance Kanamycin
Aminoglycoside-3'-phosphotransferase (APH(3')), also known as aminoglycoside kinase, is an enzyme that primarily catalyses the reaction of adding a phosphate from ATP to the 3'-hydroxyl group of a 4,6-disubstituted aminoglycoside (Figure 4)(Frontiers in Bioscience 4, d9-21, 1999). In this case kanamycin. More specific we used aph(3’)-Ia. It provides resistance against kanamycin as well as neomycin and pradimicin (Shaw, 1993).
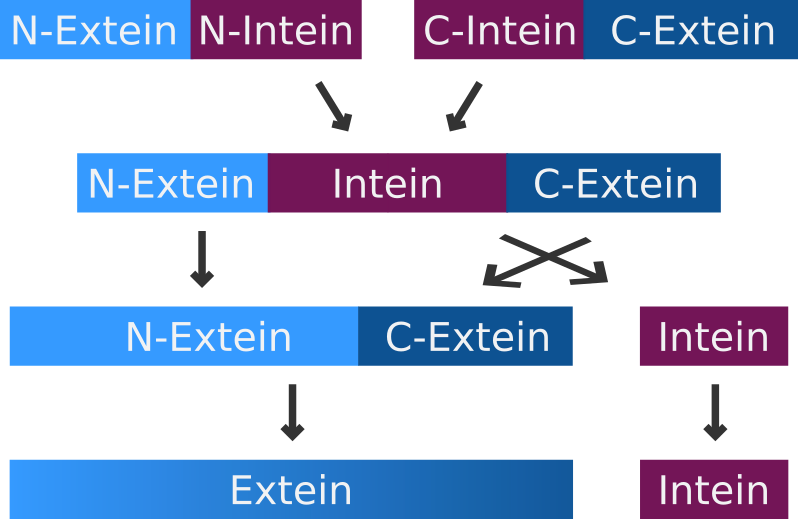
Intein-Mediated Protein Splicing
Inteins
Mechanism of intein-mediated protein splicing.
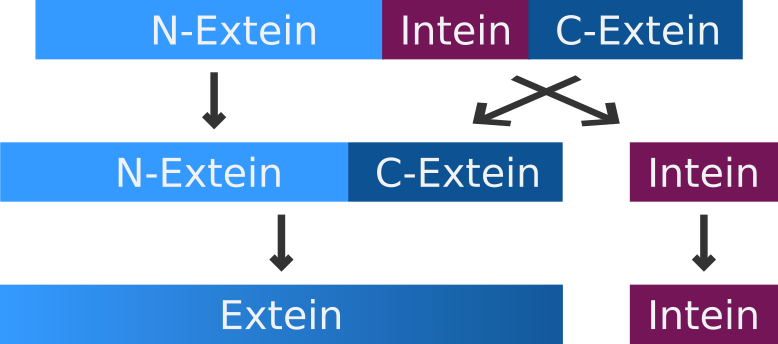
An intein is a segment of a precursor protein which is able to mediate its own excision form the precursor protein, while joining the flanking protein sequences together, creating a new peptide bond.
The intein-mediated protein splicing occurs after the mRNA is translated into a sequence of amino acids. Prior to the splicing process the N-terminal part of the precursor protein is called N-extein, the center part is the intein and the C-terminal part is named C-extein. The spliced protein is called an extein as well.
Split Inteins
A very special but small subset of inteins are the so-called split-inteins. They are transcribed and translated as separate polypeptides but rapidly associate afterwards to from the active intein. The active intein then ligates the fused N- and C-exteins in a process called protein trans-splicing.
Mechanism of split intein-mediated protein splicing.
After the first split intein SspDnaE, a subunit of the DNA polymerase III (DnaE) from Synechocystis sp. strain PCC6803 was discovered. Several homologous inteins were identified in cyanobacteria. (Wei et al. 2006, Dassa et al. 2007) However apart from sequence analysis there have only been few attempts to further characterize or compare their splicing efficiencies (Dassa et al. 2007). Nevertheless a split intein from the cyanobacterium Nostoc punctiforme (NpuDnaE) which is highly homologous to Ssp DnaE was analyzed quite extensively. It has shown higher activities than SspDnaE in vivo and in vitro (Iwai et al. 2006; Zettler et al. 2009). Additionally it has a boarder range of acceptance when it comes to the residues at the end of the C-extein (Iwai et al. 2006).
Mechanism
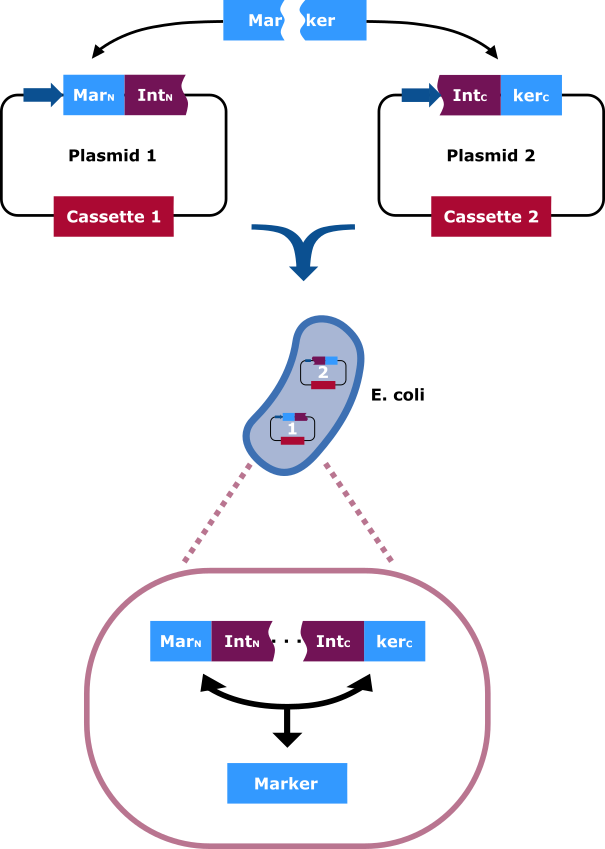
Split-Resistance Genes
The selection marker is split in two halfs. Each part of it is cloned into a different vector carrying distinct cassettes (Figure 4). The marekers N-terminal (MarN) and C-terminal (MarC) fragment is fused downstream and upstream respectively with the N-terminal (IntN) and C-terminal (IntC) fragment of the split Npu DnaE Intein. Only cells that contain both plasmids can grow on hygromycin rich medium. This occurs due to the intein fragments, because after the fusions being separately expressed, the split intein sections find one another and through trans splicing cut themselves out and fuse their flanking sites via peptide bond. Thereby an intact hygromycin-B 4-O-kinase is released and the cells gain the selection ability.
The selection marker is split in two halfs. The N-terminal and C-terminal fragment if it is fused upstream and downstream respectively of the N-terminal and C-terminal fragment of the Npu DnaE intein. Each fusion is cloned in a different backbone, so that the oris are compatible. On the protein level the split intein fragments find to each other and through protein trans-splicing they cut themselves out. Thereby an active hygromycin-B 4-O-kinase is assembled and the cells can grow on hygromycin B rich medium.
Fusion-Protein Structure
There are many important features, that must be taken into consideration while planning to construct a two-plasmid system with split antibiotics. Otherwise the probability of success decreases. While trying to find the right position to split the resistance gene it must be searched for the right N- and C-terminal extein sequences. In our case we talked to Mr. Albert Cheng, PhD who suggested us to orientate ourselves on Table 1 (Cheriyan, 2013). On this table the relative favourability of each amino acid at each relevant position neighbouring the inteins is documented. Additionally, it is important to make sure that the split point is not interrupting an important secondary structure of the protein and that the exteins are not hydrophobic. Thanks to Dr Francine Perler reminding us about the hydrophobicity of the exteins, we did not forget to take it into consideration. Instead of analysing ourselves the DNA sequence of the resistance genes we decided to do a model, that will be able to output the right split points.
As a positive reference to our selected split points we contacted Prof. Dr. Barbara Di Ventura. She provided us with the plasmids that contain the protein fusions, so that we can use them to clone our new backbones. Two plasmids encoding an active TEM-1 lactamase for ampicillin resistance and other two plasmids encoding the aph(3’)-Ia for kanamycin resistance (not published jet, Navaneethan Palanisam, Freiburg University).
Relative favorability SrF) of each amino acid at each relevant position neighboring the inteins. The favorability of each amino acid in each position, -1, -2, -3 for N-terminal extein and -+1, +2, +3 for C-terminal extein is listed. Adjusted from Cheriyan, Pedamallu, Tori, & Perler, 2013.
Amino Acid
rF at the N-terminal extein
rF at the C-terminal extein
-3
-2
-1
+1
+2
+3
D
1.39
1.52
0.00
0.00
0.00
1.26
E
1.26
3.51
0.07
0.00
0.00
5.5
N
0.93
1.26
2.19
0.00
0.00
8.61
Q
1.39
0.86
0.33
0.00
0.00
0.27
H
1.06
1.26
2.19
0.00
0.00
8.61
K
1.59
0.93
4.44
0.00
0.07
0.00
R
1.81
0.2
0.86
0.00
0.00
0.00
S
0.99
1.24
1.37
0.00
0.00
0.07
C
0.46
0.40
0.80
50
0.07
0.07
T
0.63
0.89
1.59
0.00
0.00
0.96
P
0.80
1.52
0.00
0.00
0.00
0.00
G
4.67
2.68
0.17
0.00
0.00
0.03
A
0.63
0.83
1.92
0.00
0.00
0.00
V
0.56
0.50
0.23
0.00
0.03
0.10
I
0.07
0.53
0.00
0.00
0.00
0.40
L
0.11
0.57
0.75
0.00
0.00
0.82
M
0.13
0.73
1.26
20
0.07
2.25
F
0.13
0.73
1.26
0.00
0.07
2.25
Y
0.20
0.60
2.05
0.00
0.13
5.23
W
0.07
0.40
0.99
0.00
29.42
0.07
Characterization
Single Plasmid Control
An important aspect, when constructing and working with a split-resistance gene system is to make sure, that one plasmid does not provide to the cells the ability to grow on medium with the corresponding antibiotic. Therefore, from every cell collection containing one of the plasmid pairs, we plated different on plates with low antibiotic concentration.
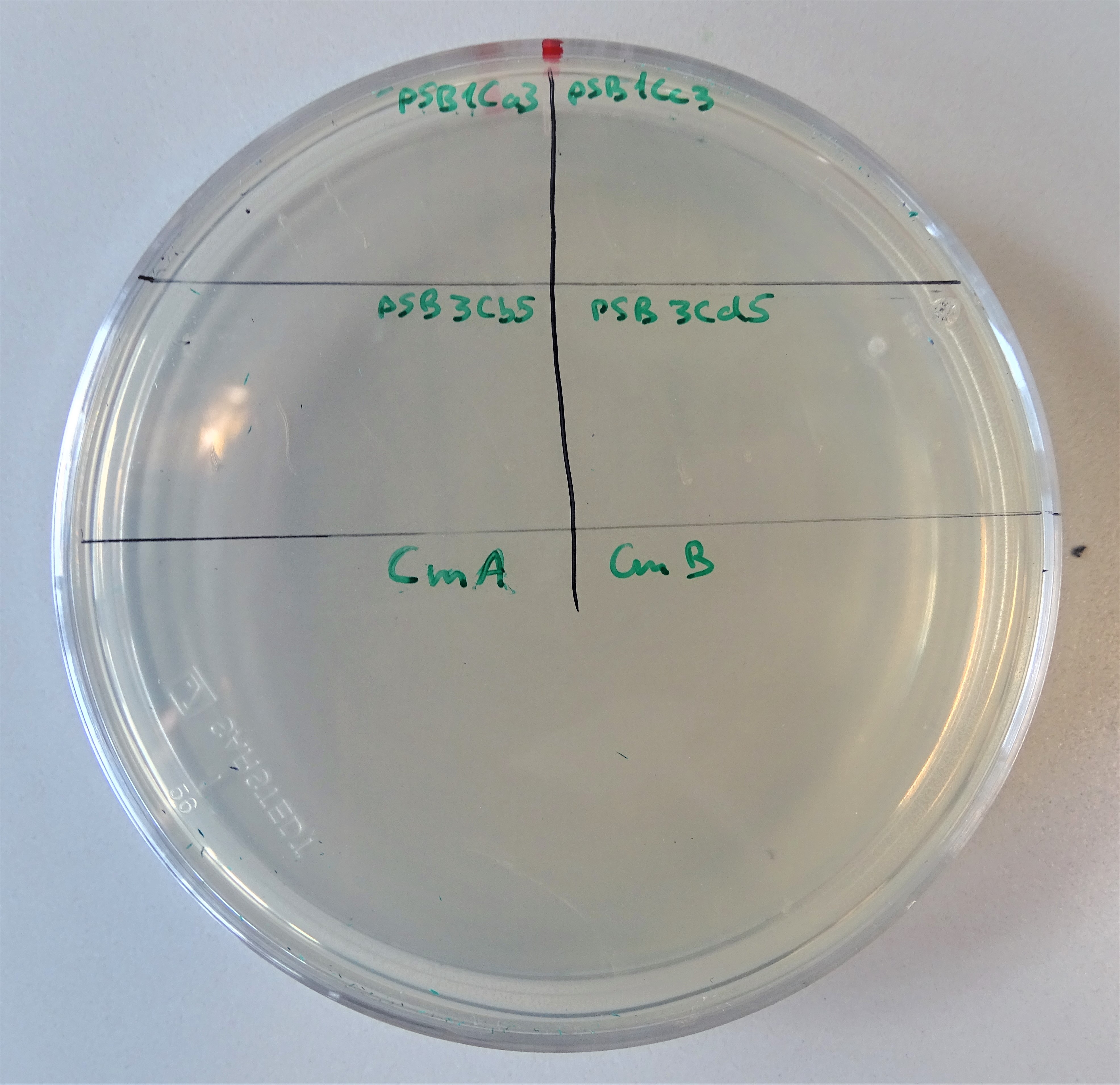
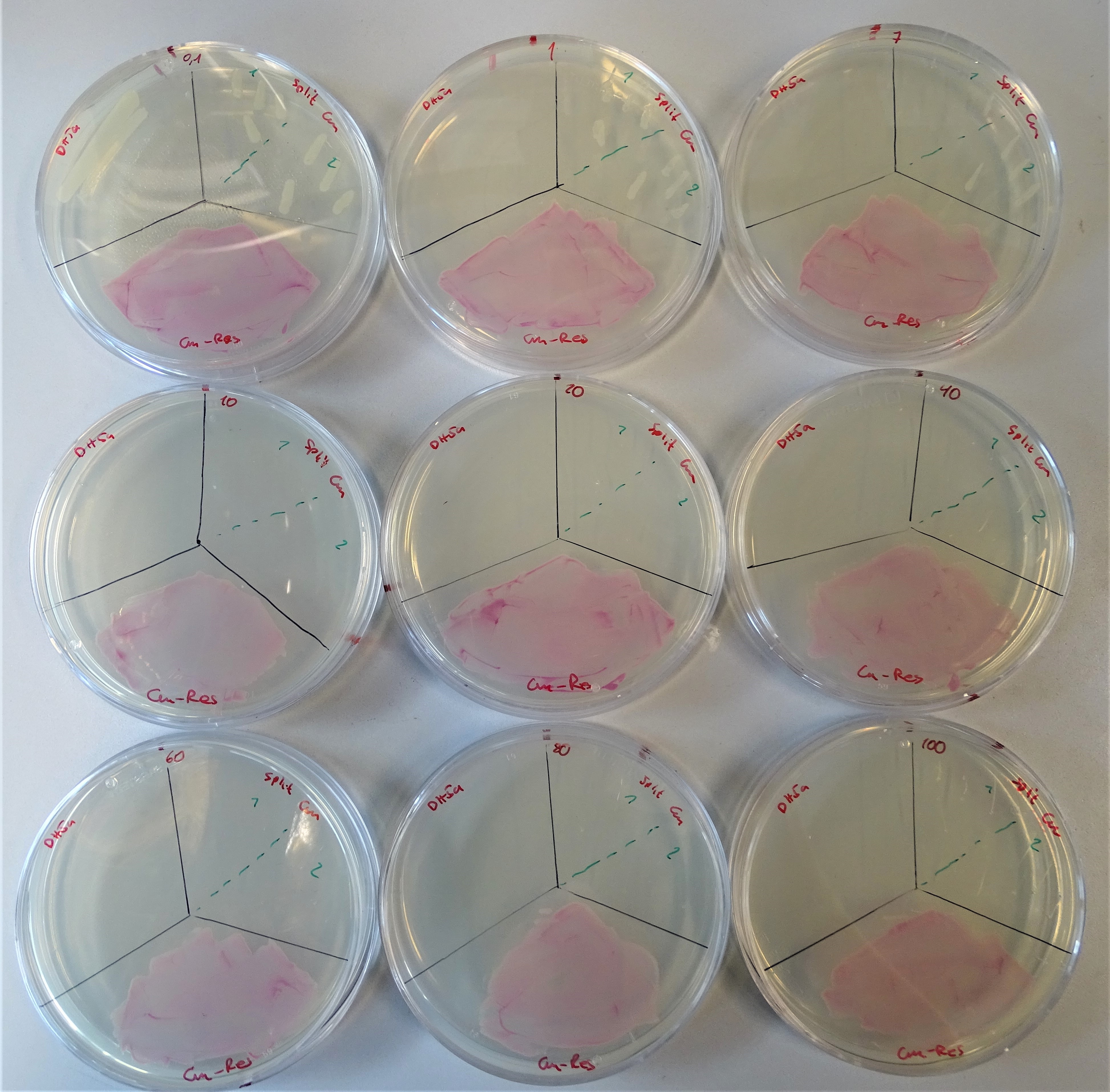
Normal working concentrations for chloramphenicol are between 25 and 170 µg/mL (ATCC_recommendations_antibiotic_concentrations, 2003). As Figure 11 shows the chloramphenicol plate was divided into four segments. Cells were plated on the upper left site containing pSB1Ca3, upper right site containing pSB1Cc3, lower left site containing pSB3Cb5 and on the lower right site containing pSB3Cd5. On none of the segments growing colonies can be seen and that proves, that only one plasmid does not provide antibiotic resistance. Similar is the case with hygromycin.
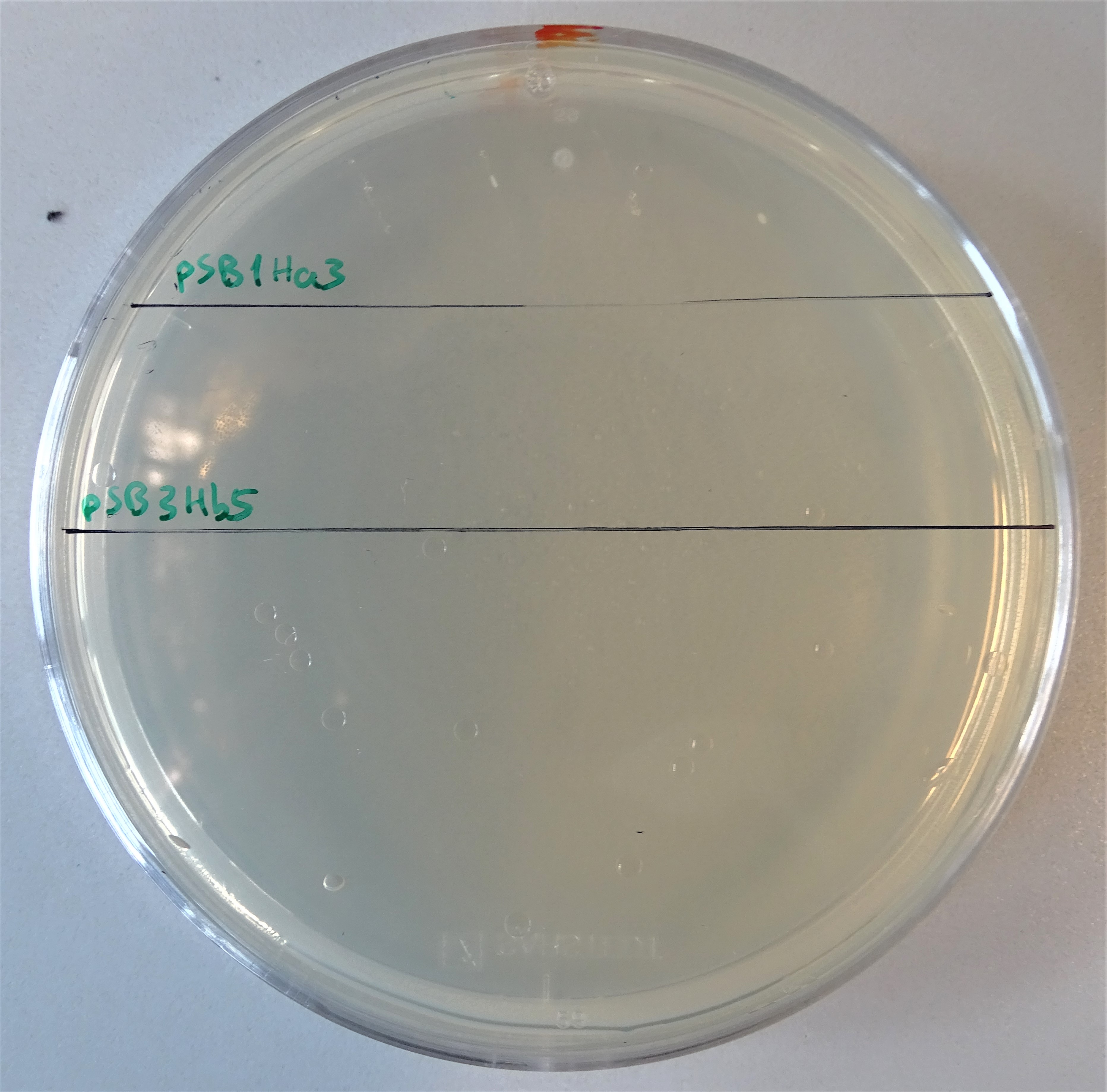
Normal working concentrations for hygromycin B is between 200 and 500 µg/mL (Hygromycin B, Thermofisher). As Figure 12 shows the hygromycin plate was divided into two segments. Cells were plated on the upper site containing pSB1Ha3 and on the lower site containing pSB3Hb5. Like before, none of the plated colonies grew.
Plate with a chloramphenicol concentration of 5 μg/mL. Cells were plated containing, on the upper left site pSB1Ca3, upper right site pSB1Cc3, lower left site pSB3Cb5 and on the lower right site pSB3Cd5
Plate with a hygromycin B concentration of 50 μg/mL. On the upper half, cells were plated that contain only the pSB1Ha3 plasmid. On the lower half, cells were plated that contain only the pSB3Hb3 plasmid.
Growth Control
After proving that one plasmid does not provide an antibiotic resistance, the next experiment was to analyse the growth efficiency of the double-plasmid containing cells. Therefore, we poured plates with diverse concentrations of chloramphenicol and hygromycin B (Figure 13, 14).
Regarding the chloramphenicol resistance the concentrations that were prepared are from the upper left to the lower right site 0.1, 1, 7, 10, 20, 40, 60, 80, 100 μg/mL. Each plate was divided into three sections. On the upper left site, DH5α was plated as the negative control, on the upper right site the colonies containing both plasmids and on the lower site a colony containing pSB1C3 (link) gene. The maximal chloramphenicol concentration on which DH5α grows is 0.1 μg/mL. The section for the split-marker was additionally divided into two segments, for the two different split point of chloramphenicol. On the left part the CmA version and on the right part the CmB version was plated. Until a concentration of 1 µg/mL and in other two repeats till a concentration of 5 µg/mL the two split system versions were providing the cell resistance against chloramphenicol. Furthermore, it can be observed from the figure that the CmB version works better than the other one. This was to assume, because of the extein sequence flanking the split Npu DnaE intein.
From the upper left site to, to the right till the lower right site, plates are lined up with the hygromycin concentrations 40, 30, 60 80, 100, 200, 500, 1000μg/mL. Every plate is divided into three sections. On the upper left site DH5α was plated representing the negative control, on the upper right site colonies containing both plasmids and on the lower site a colony containing one plasmid with a hygromycin resistance gene.
From the upper left site to, to the right till the lower right site, plates are lined up with the hygromycin concentrations 30, 40, 60 80, 100, 200, 500, 1000μg/mL. Every plate is divided into three sections. On the upper left site DH5α was plated representing the negative control, on the upper right site colonies containing both plasmids and on the lower site a colony containing one plasmid with a hygromycin resistance gene.
For the hygromycin B resistance the concentrations that were prepared are from the upper left to the lower right site 30, 40, 60 80, 100, 200, 500 and 1000 μg/mL. Each plate was divided into three sections. On the upper left site, DH5α was plated as the negative control, on the upper right site the colonies containing both plasmids and on the lower site a colony containing plasmid with a hygromycin resistance gene. For DH5α the used concentrations were too high to grow. The positive control did not have any problems growing on hygromycin B. The cells with the split antibiotic resistance plasmids easily grew on the plates till a concentration of 500 μg/mL and with some difficulties on 1000 μg/mL.
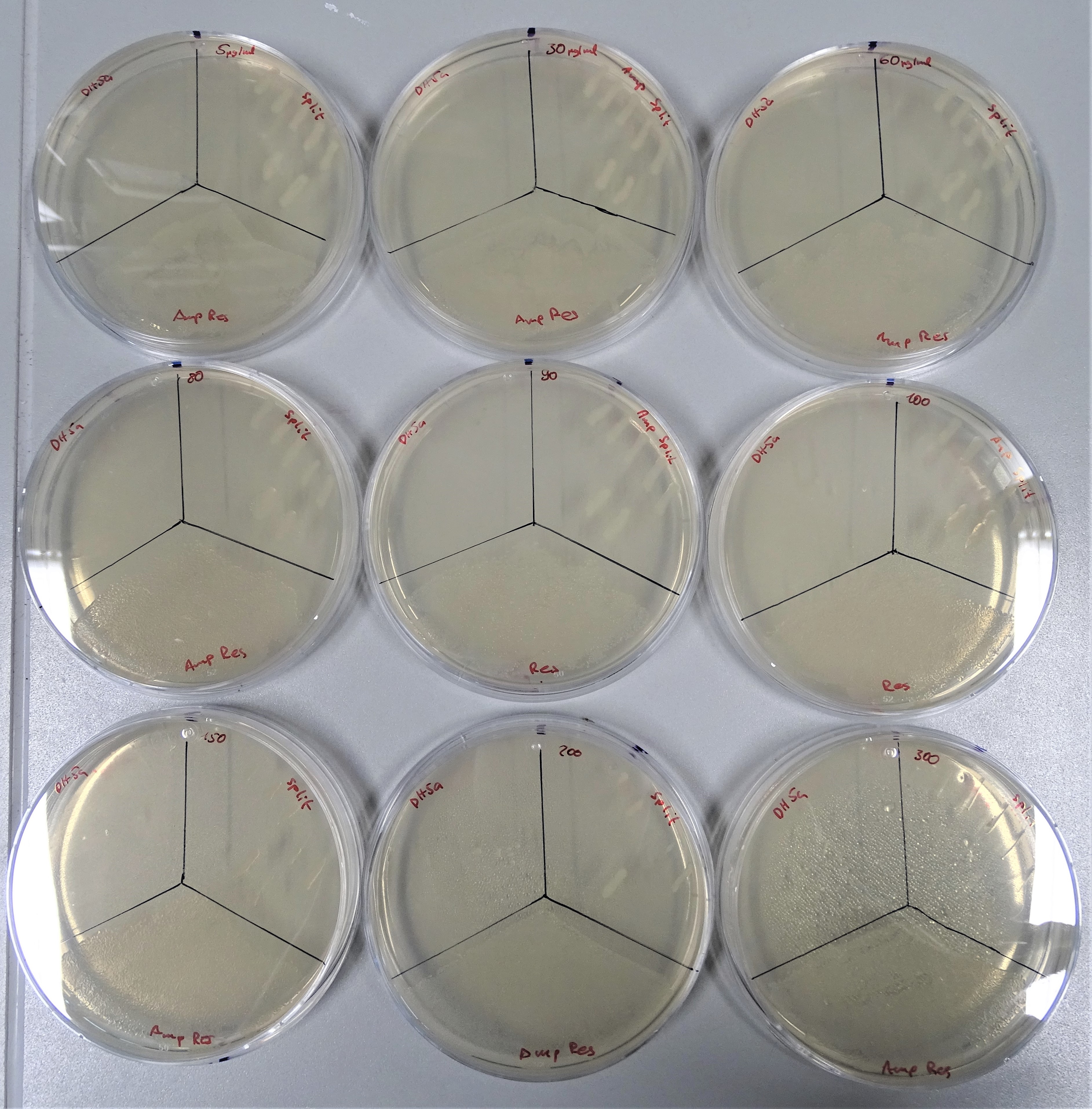
Regarding the ampicillin resistance the concentrations that were prepared are from the upper left to the lower right site 5, 30, 60, 80, 90, 100, 150, 200 and 300 μg/mL (Fig.16). The plate was divided into three sections. On the upper left site, DH5α was plated as the negative control, on the upper right site the colonies containing both plasmids and on the lower site a colony containing pTXB1. DH5α could not grow on any of these concentrations. The positive control was also able to grow on any ampicillin concentration. The cells with the split antibiotic resistance plasmids easily grew on the plates till a concentration of 200 μg/mL.
dcfvgh
dcfvgh
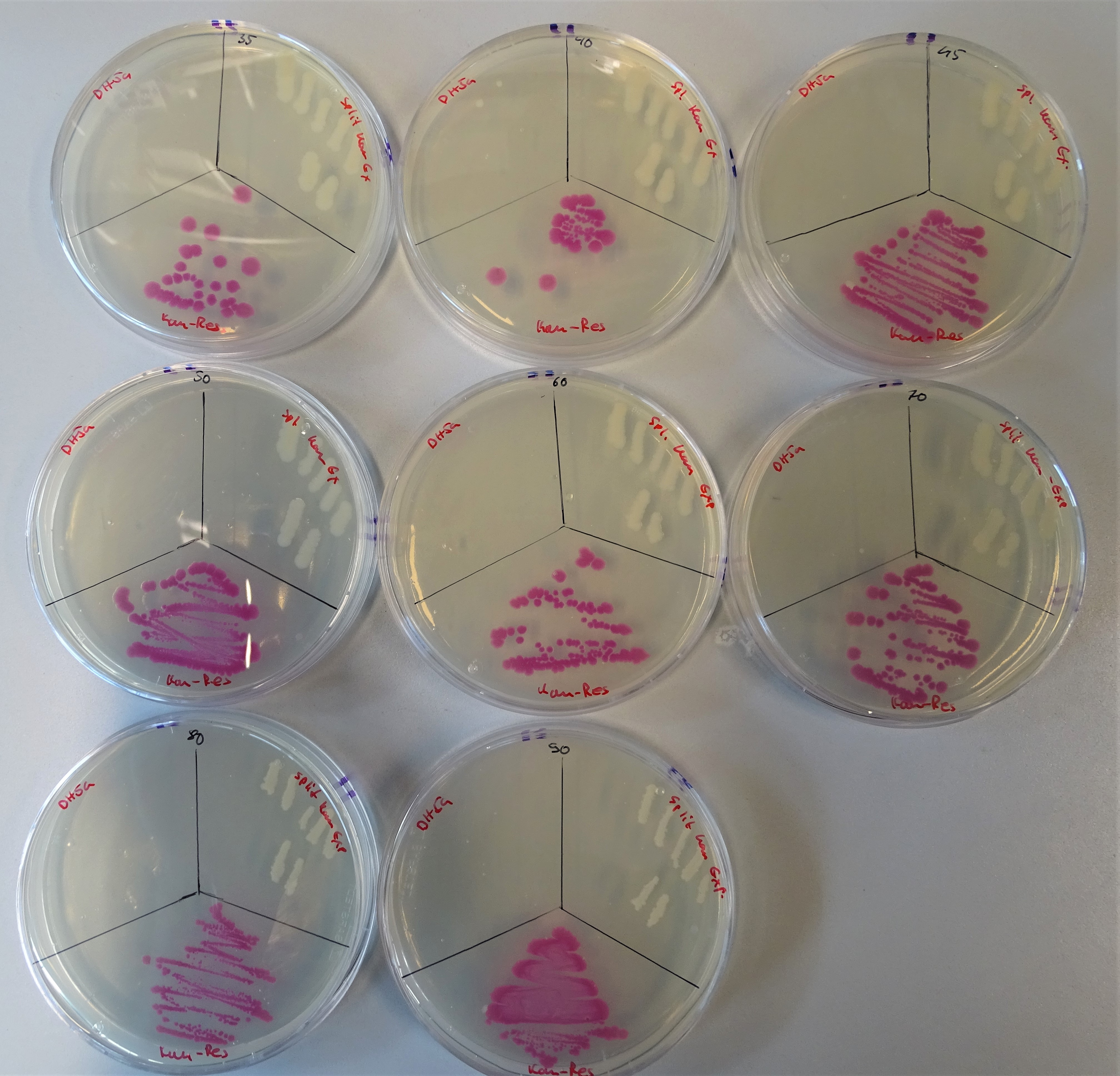
For the kanamycin resistance the concentrations that were prepared are from the upper left to the lower right site 35, 40, 45, 50, 60, 70, 80 and 90μg/mL (Fig.17). As before too, the plate was divided into three sections. On the upper left site, DH5α was plated as the negative control, on the upper right site the colonies containing both plasmids and on the lower site a colony containing pSB1K3. For DH5α the used concentrations were too high to grow, except of the 40µg/ml plate, where two colonies can be seen. The positive control was also able to grow on any ampicillin concentration. The cells with the split antibiotic resistance plasmids easily grew on the plates till a concentration of 90 μg/mL.
Growth Control in LB-Medium
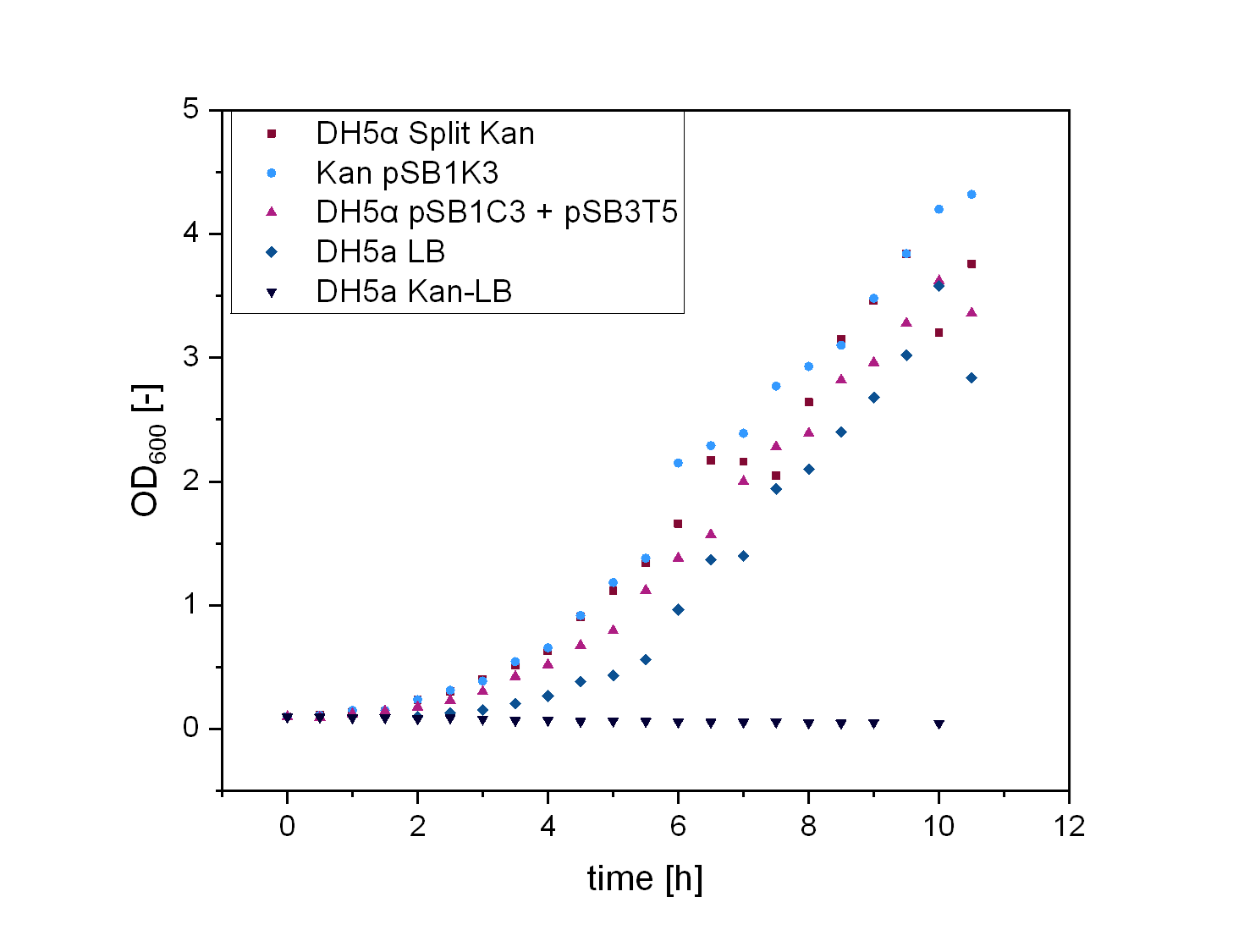
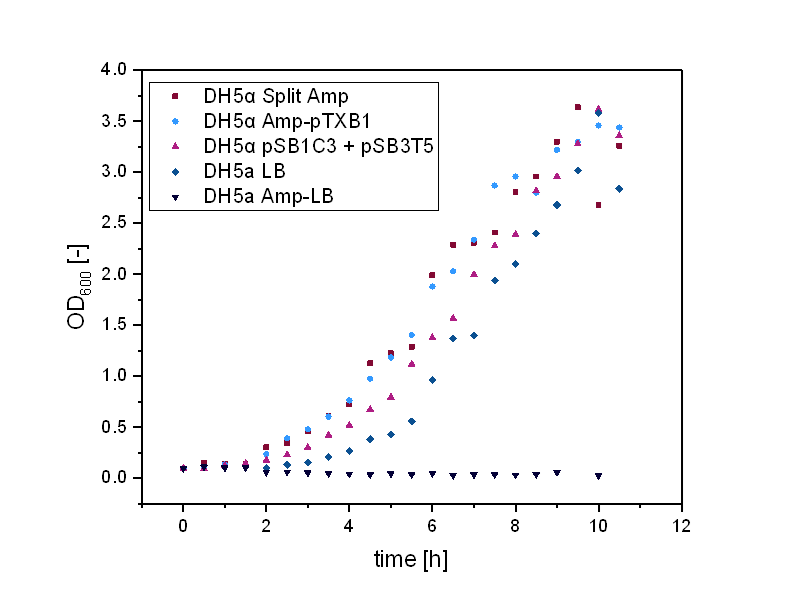
In order to test the functionality of our split resistance genes, we conducted growth experiments with DH5α containing the two-plasmid system. The Escherichia colis (E. colis) were cultivated in liquid LB media and all tested plasmids were transformed in DH5α. For every growth curve there were five samples. For the split kanamycin curve the first sample was DH5α cultivated in LB-media without antibiotic, the second DH5α in LB-kanamycin, the third DH5α + pSB1K3 in LB-kanamycin, the fourth DH5α + pSB1C3 + pSB3T5 in LB-kanamycin-tetracyclin, the fifth DH5α + pSB1Ka3 + pSB3Kb5 in LB-kanamycin.
DH5α with antibiotic did not grow in comparison to DH5α without antibiotic that showed the expected curve. The cells containing pSB1K3 grew the best and the split antibiotic system followed . It could also be observed, that the cells containing two plasmids with each one antibiotic resistance did grow less efficient than the split kanamycin cells.
Growth experiment of Escherichia colis (E. colis) . DH5α + pSB1Ka3 + pSB3Kb5, DH5α + pSB1K3, DH5α + pSB1C3 + pSB3T5, DH5α in LB-medium with and without kanamycin (illustrated by squares, circles, triangles, reverse triangles and diamonds). The cells were grown at a temperature of 30 degrees Celsius at 180 rpm.
The same procedure was conducted to test the functionality of the split ampicillin system. The Escherichia colis (E. colis) were also cultivated in liquid LB media and all tested plasmids were transformed in DH5α. The first sample was DH5α cultivated in LB-media without antibiotic, the second DH5α in LB-kanamycin, the third DH5α + pTXB1 in LB-ampicillin, the fourth DH5α + pSB1C3 + pSB3T5 in LB-chloramphenicol-tetracyclin, the fifth DH5α + pSB1Aa3 + pSB3Ab5 also in LB-ampicillin.
DH5α with antibiotic did not grow in comparison to DH5α without antibiotic, as also seen before. The cells containing pSB1K3 grew as good as the cells with the split antibiotic system. DH5α with two antibiotic resistance genes did grow better than DH5α with antibiotic, but worser than the others.
Growth experiment of Escherichia colis (E. colis) . DH5α + pSB1Ka3 + pSB3Kb5, DH5α + pTXB1, DH5α + pSB1C3 + pSB3T5, DH5α in LB-medium with and without kanamycin (illustrated by squares, circles, triangles, reverse triangles and diamonds). The cells were grown at a temperature of 30 degrees Celsius at 180 rpm.
Modeling Split Chloramphenicol Resistance
We aimed to split the protein conveying resistance to chloramphenicol, chloramphenicol acetyltransferase (BBa_J3105). The resistance protein is commonly used within iGEM, thus making a split Chloramphenicol acetyltransferasa a vluable addition to the iGEM partsreg. The protein sequence of this part can be found here.
According to our modeling, the best split point to use when introducing Npu DnaE was VAQ-CTY in the positions 28-33, the B(VAQCTY) is 56.95.
Even though this split point is located at the second amino acid of a beta-sheet, we decided to test whether splitting at this position could lead to Chloramphenicol acetyltransferase fragments that could be reconstituted by intein-mediated protein splicing.
Similar to our modeling of the split kanamycin resistance, we used the homology modeling software MODELLER (Sali & Webb, 1989) to visualize each part of the chloramphenicol acetyltransferase fused to the Npu DnaE intein.
The amino acid sequences given to the software were:
Part 1:
MEKKITGYTTVDISQWHRKEHFEAFQSVAQCLSYETEILTVEYGLLPIGKIVEKRIECTVYSVDNNGNIYTQPVAQWHDRGEQEVFEYCLEDGSLIRATKDHKFMTVDGQMLPIDEIFERELDLMRVDNLPNNLEGHHHHHH
Part 2:
MIKIATRKYLGKQNVYDIGVERDHNFALKNGFIASNCCTYNQTVQLDITAFLKTVKKNKHKFYPAFIHILARLMNAHPEFRMAMKDGELVIWDSVHPCYTVFHEQTETFSSLWSEYHDDFRQFLHIYSQDVACYGENLAYFPKGFIENMFFVSANPWVSFTSFDLNVANMDNFFAPVFTMGKYYTQGDKVLMPLAIQVHHAVCDGFHVGRMLNELQQYCDEWQGGA
The templates used for homology modeling were:
Part 1
Part 2
1NOC, chain B (Chloramphenicol acetyltransferase)
1NOC, chain B (Chloramphenicol acetyltransferase)
4QFQ, chain A (Npu DnaE)
4QFQ, chain B (Npu DnaE)
5OL6, chain A (inactivated Npu SICLOPPS intein with CAFHPQ extein)
5OL6, chain B (inactivated Npu SICLOPPS intein with CAFHPQ extein)
4KL5, chain A (Npu DnaE)
1QCA, chain A (Type III Chloramphenicol acetyltransferase)
2KEQ, chain A (DnaE intein from Nostoc punctiforme)
3CLA, chain A (Type III Chloramphenicosl acetyltransferase)
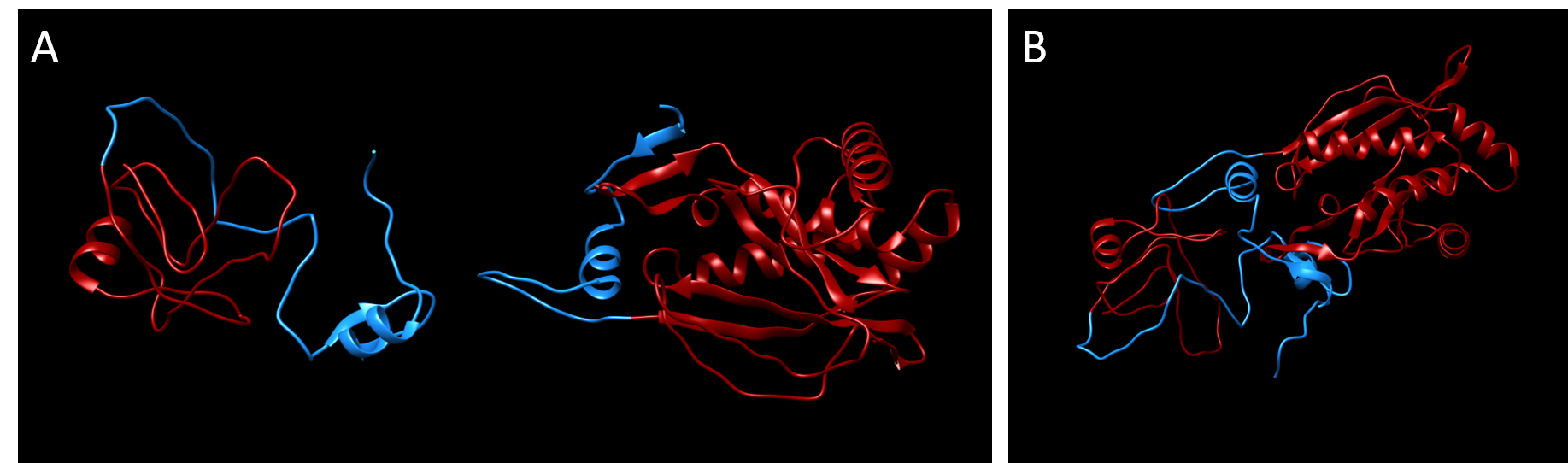
The predicted 3D-structure of each subunit of the Cloramphenicol acetyltransferase split at the optimal predicted split point VAQ-CTY prior to splicing. It was developed using the homology modeling software MODELLER (Sali & Webb, 1989).
As shown in growth experiments on plates (Fig. 9), implementing split antibiotic resistances is feasible. Using the split points predicted with the model allowed us to successfully construct functional split antibiotic resistances.
Modeling Split Kanamycin Resistance
We aimed to split the protein conveying resistance to chloramphenicol, chloramphenicol acetyltransferase (BBa_J3105). The resistance protein is commonly used within iGEM, thus making a split Chloramphenicol acetyltransferasa a vluable addition to the iGEM partsreg. The protein sequence of this part can be found here.
According to our modeling, the best split point to use when introducing Npu DnaE was VAQ-CTY in the positions 28-33, the B(VAQCTY) is 56.95.
Even though this split point is located at the second amino acid of a beta-sheet, we decided to test whether splitting at this position could lead to Chloramphenicol acetyltransferase fragments that could be reconstituted by intein-mediated protein splicing.
Similar to our modeling of the split kanamycin resistance, we used the homology modeling software MODELLER (Sali & Webb, 1989) to visualize each part of the chloramphenicol acetyltransferase fused to the Npu DnaE intein.
The amino acid sequences given to the software were:
Part 1:
MEKKITGYTTVDISQWHRKEHFEAFQSVAQCLSYETEILTVEYGLLPIGKIVEKRIECTVYSVDNNGNIYTQPVAQWHDRGEQEVFEYCLEDGSLIRATKDHKFMTVDGQMLPIDEIFERELDLMRVDNLPNNLEGHHHHHH
Part 2:
MIKIATRKYLGKQNVYDIGVERDHNFALKNGFIASNCCTYNQTVQLDITAFLKTVKKNKHKFYPAFIHILARLMNAHPEFRMAMKDGELVIWDSVHPCYTVFHEQTETFSSLWSEYHDDFRQFLHIYSQDVACYGENLAYFPKGFIENMFFVSANPWVSFTSFDLNVANMDNFFAPVFTMGKYYTQGDKVLMPLAIQVHHAVCDGFHVGRMLNELQQYCDEWQGGA
The templates used for homology modeling were:
Part 1
Part 2
1NOC, chain B (Chloramphenicol acetyltransferase)
1NOC, chain B (Chloramphenicol acetyltransferase)
4QFQ, chain A (Npu DnaE)
4QFQ, chain B (Npu DnaE)
5OL6, chain A (inactivated Npu SICLOPPS intein with CAFHPQ extein)
5OL6, chain B (inactivated Npu SICLOPPS intein with CAFHPQ extein)
4KL5, chain A (Npu DnaE)
1QCA, chain A (Type III Chloramphenicol acetyltransferase)
2KEQ, chain A (DnaE intein from Nostoc punctiforme)
3CLA, chain A (Type III Chloramphenicosl acetyltransferase)
After running the MODELLER software (Sali & Webb, 1989), two protein structures were calculated and placed adjacent to each other using Chimera (Pettersen et al., 2004).
The predicted 3D-structure of each subunit of the Cloramphenicol acetyltransferase split at the optimal predicted split point VAQ-CTY prior to splicing. It was developed using the homology modeling software MODELLER (Sali & Webb, 1989).
As shown in growth experiments on plates (Fig. 9), implementing split antibiotic resistances is feasible. Using the split points predicted with the model allowed us to successfully construct functional split antibiotic resistances.