
Difference between revisions of "Part:BBa K1028002"
(→Introduction) |
(→Introduction) |
||
| (16 intermediate revisions by the same user not shown) | |||
| Line 5: | Line 5: | ||
<img src="http://i276.photobucket.com/albums/kk29/JosephBeetBeet/BS1withnotshitonit.png" style="width:600px;margin-right:160px" ></a> | <img src="http://i276.photobucket.com/albums/kk29/JosephBeetBeet/BS1withnotshitonit.png" style="width:600px;margin-right:160px" ></a> | ||
| − | <p style="text-align:center"> | + | <p style="text-align:center"> |
</p></html> | </p></html> | ||
| − | The CpxR promoter is | + | This BioBrick is designed to act as a reporter for hydrophobic membrane stress via surface binding of cells to macro-structures in solution, utilising the CpxR promoter and GFP to act as a reporter. The CpxR promoter is know to react to hydrophobic membrane stress, something that can be imposed on a cell via binding with large structures. This does give the Part a limitation in that any binding on the molecular scale will go unnoticed by the promoter. |
| + | |||
| + | ===General Cpx Information=== | ||
| + | <html> | ||
| + | <img src="http://i276.photobucket.com/albums/kk29/JosephBeetBeet/cpxshcematiSIMPLFIED.png" style="width:400px;margin-right:160px" ></a> | ||
| + | |||
| + | <p style="text-align:center"><b>Fig.1:</b> Illustrated schematic of the Cpx system in full, showing the multitude of protein interactions involved in relaying the signal to create transcriptional activation. | ||
| + | </p></html> | ||
| + | |||
| + | This part is highly dependent upon a naturally occuring pathway in E.Coli called the Cpx pathway. It is associated with the regulation of periplasmic membrane stress and the misfolding of surface proteins. It is a two component signal transduction pathway. The first component is CpxA, an inner membrane protein which, when bound to CpxP (an inhibitor molecule), in inactive. When CpxP is unbound from CpxA, CpxA autophosphorylates itself. A kinase enzyme then phosphorylates the CpxR, the second component in the pathway, which then binds to the Cpx promoter and activates transcription. There is still a lot of debate regarding this pathway and many different theories but it is thought that membrane stress causes the misfolding of proteins (pili being the main contributor) causes misfolded subunits to accumulate in the periplasm and titrate the CpxP inhibitor molecule off the CpxA and activate the pathway. | ||
| + | |||
| + | Increasing the specificity of the CpxR promoter for hydrophobic membrane stress using methods such as intentionally introducing mutations in the promoter.In addition to this utilisation of an off-switch regulator, CpxP, to additonal pathways used to create a bio-logic gate or even by optimisation of buffer solution could be utilized to achieve this added specificity. | ||
==='''Experimental Data'''=== | ==='''Experimental Data'''=== | ||
| Line 31: | Line 42: | ||
</p> | </p> | ||
====Triton X-100==== | ====Triton X-100==== | ||
| − | " | + | <html> |
| − | < | + | <img src="https://static.igem.org/mediawiki/2013/thumb/0/0a/Triton_bs1_graph-no_line_of_best_fit.png/400px-Triton_bs1_graph-no_line_of_best_fit.png" style="width:450px;margin-left:160px" ></a> |
| + | |||
| + | <p style="text-align:center"><b>Fig.1: </b> Testing the effect of the detergent Triton X-100 on the CpxR promoter | ||
| + | </p> | ||
| + | <p style="text-align:left"> | ||
Between the concentration values of 0.1-9% triton X-100 the data shows roughly the relationship that was expected, a rise in GFP production as the triton concentration increases. This is likely due to triton acting as a detergent resulting in damage to the plasma membrane of cells, activating the Cpx pathway in general, including the cpxR promoter. | Between the concentration values of 0.1-9% triton X-100 the data shows roughly the relationship that was expected, a rise in GFP production as the triton concentration increases. This is likely due to triton acting as a detergent resulting in damage to the plasma membrane of cells, activating the Cpx pathway in general, including the cpxR promoter. | ||
Past 10% Triton X-100 concentration it is assumed that cells are dying in large quantities, being unable to produce GFP at all or in very limited amounts before they are killed. This explains the corrected fluorescence readings reducing to below zero. | Past 10% Triton X-100 concentration it is assumed that cells are dying in large quantities, being unable to produce GFP at all or in very limited amounts before they are killed. This explains the corrected fluorescence readings reducing to below zero. | ||
| − | The reason for the large spike in fluorescence at Triton concentrations >20% is unclear, a possibly explanation for this is the total lysis of the cell, expelling all cellular components resulting in background GFP levels having a larger fluorescence reading due to reduced scattering from within the cell | + | The reason for the large spike in fluorescence at Triton concentrations >20% is unclear, a possibly explanation for this is the total lysis of the cell, expelling all cellular components resulting in background GFP levels having a larger fluorescence reading due to reduced scattering from within the cell |
| + | </p> | ||
| + | </html> | ||
====pH Effects==== | ====pH Effects==== | ||
| − | http://i276.photobucket.com/albums/kk29/JosephBeetBeet/phbs1graph.png | + | <html> |
| + | <img src="http://i276.photobucket.com/albums/kk29/JosephBeetBeet/phbs1graph.png" style="width:450px;margin-left:160px" ></a> | ||
| + | |||
| + | <p style="text-align:center"><b>Fig.3:</b> The effects of a range of pH values on the activation of the CpxR promoter | ||
| + | </p></html> | ||
| + | |||
<br> | <br> | ||
The slight fluctuations across the pH scale are in line with the background fluorescence readings from our cell cultures, representing the standard levels of cpx activation as the cells collide, disperse in solution and divide during replication. | The slight fluctuations across the pH scale are in line with the background fluorescence readings from our cell cultures, representing the standard levels of cpx activation as the cells collide, disperse in solution and divide during replication. | ||
The higher levels of expression at both low and high pH are likely to be due to the pH extremes denaturing proteins and disrupting interactions within the plasma membrane. It could be assumed that pH’s further into the extremes of acid and base would completely lyse and kill the cells resulting in a possibly lower fluorescence. | The higher levels of expression at both low and high pH are likely to be due to the pH extremes denaturing proteins and disrupting interactions within the plasma membrane. It could be assumed that pH’s further into the extremes of acid and base would completely lyse and kill the cells resulting in a possibly lower fluorescence. | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
===Atomic Force Microscopy === | ===Atomic Force Microscopy === | ||
| Line 58: | Line 74: | ||
</p></html> | </p></html> | ||
| + | The Atomic Force Microscopy experiments were designed to quantify the force necessary to trigger the CpxR promoter, generating GFP. This was to be done by observing the culture under the fluoroscope portion of the F-AFM, and then "prod" a bacterium at a low force, slowly increasing the stress until a fluorescence response was seen. This would then give a direct measure of the stress threshold for pCpxR, a value currently unknown in the literature. However, as with all new experiments, there were some teething issues. Firstly, it can be seen from the image that the culture fluoresced at a low level prior to exposure to the AFM tip. This was likely due to smaller stresses in solution and some can clearly be seen in the middle of binomial fission, which is known to also activate the Cpx pathway in general. There were also issues with trying to keep the bacterial cells stable enough to continually poke them with the AFM tip, initially a high glycerol concentration was employed in an attempt to achieve this however the cells remained very mobile. | ||
| + | <br> | ||
| + | <br> | ||
<!-- Add more about the biology of this part here | <!-- Add more about the biology of this part here | ||
===Usage and Biology=== | ===Usage and Biology=== | ||
Latest revision as of 18:00, 4 October 2013
A cell membrane stress reporter gene
Introduction
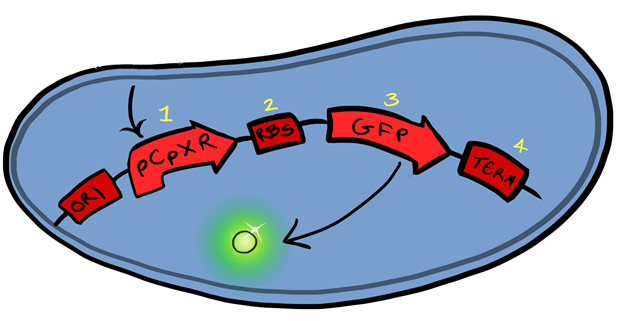
This BioBrick is designed to act as a reporter for hydrophobic membrane stress via surface binding of cells to macro-structures in solution, utilising the CpxR promoter and GFP to act as a reporter. The CpxR promoter is know to react to hydrophobic membrane stress, something that can be imposed on a cell via binding with large structures. This does give the Part a limitation in that any binding on the molecular scale will go unnoticed by the promoter.
General Cpx Information
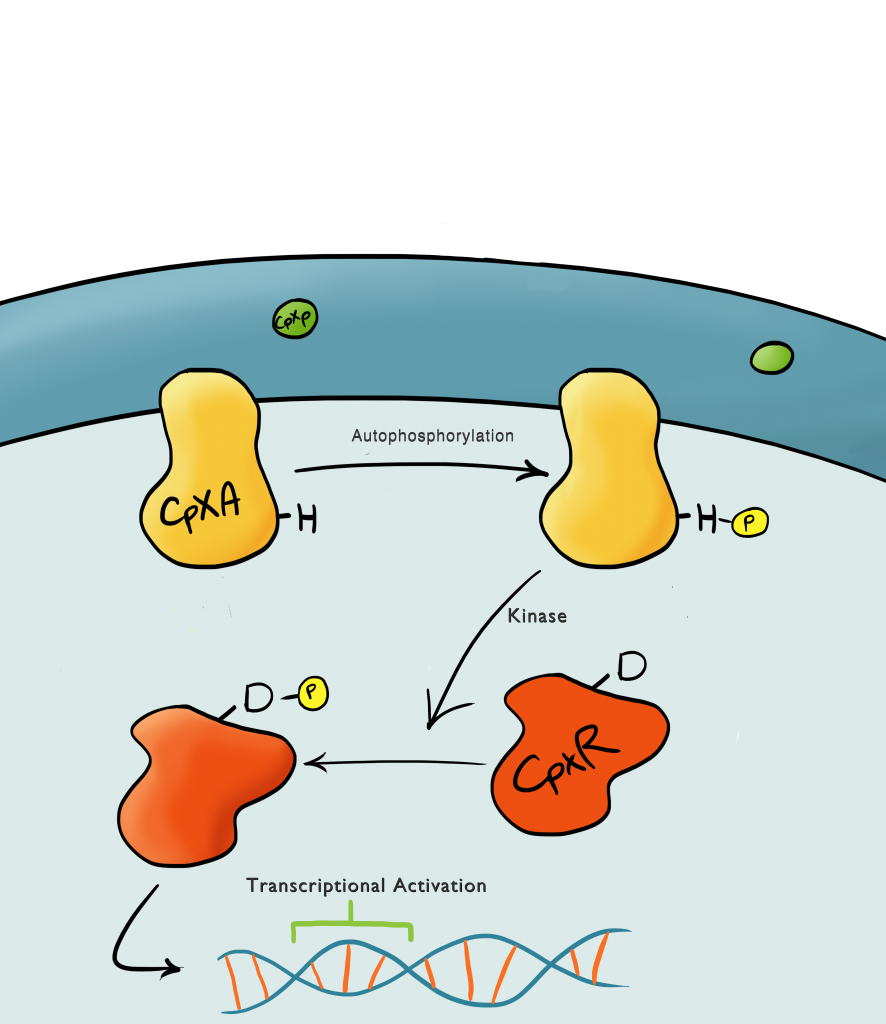
Fig.1: Illustrated schematic of the Cpx system in full, showing the multitude of protein interactions involved in relaying the signal to create transcriptional activation.
This part is highly dependent upon a naturally occuring pathway in E.Coli called the Cpx pathway. It is associated with the regulation of periplasmic membrane stress and the misfolding of surface proteins. It is a two component signal transduction pathway. The first component is CpxA, an inner membrane protein which, when bound to CpxP (an inhibitor molecule), in inactive. When CpxP is unbound from CpxA, CpxA autophosphorylates itself. A kinase enzyme then phosphorylates the CpxR, the second component in the pathway, which then binds to the Cpx promoter and activates transcription. There is still a lot of debate regarding this pathway and many different theories but it is thought that membrane stress causes the misfolding of proteins (pili being the main contributor) causes misfolded subunits to accumulate in the periplasm and titrate the CpxP inhibitor molecule off the CpxA and activate the pathway.
Increasing the specificity of the CpxR promoter for hydrophobic membrane stress using methods such as intentionally introducing mutations in the promoter.In addition to this utilisation of an off-switch regulator, CpxP, to additonal pathways used to create a bio-logic gate or even by optimisation of buffer solution could be utilized to achieve this added specificity.
Experimental Data
Investigation of literature regarding the action and regulation of the Cpx pathway lead to the discovery that the system as a whole responds to a large variety of membrane stresses, something that could be true of the specific promoter we chose to use (pCpxR).
As a result of this we deemed it necessary to investigate the response of our promoter to each individual membrane stress.
http://i276.photobucket.com/albums/kk29/JosephBeetBeet/BS1CharacterisationBars.png
Individually many of the experiments produced erratic and seemingly random data which is listed independently below. However plotting all average readings together shows that there is a dramatic increase in response to the stress induced by the silica beads comparative to all the chemically induced membrane stresses we used.
This indicates that it would be possible to detect the presence of silica beads in solution, in principle, using our binding and detection system, certainly when silica beads are present at relatively high concentrations. However further work needs to be done to assess whether this high response remains across a range of silica bead concentrations in solution.
In light of these results, some of these experiments were repeated for more accurate results. How the Cpx pathway reacts to temperature is already well characterised in the literature and by the [http://2010.igem.org/Team:Calgary/Parts/Characterization Calgary 2010 iGem team.]
The Ethanol gradient, Triton X-100 gradient and pH gradient were all repeated. In the second experiment, a robotic pipette was used to directly fill the 96 well plate to reduce human error. Excluding wells with imposed pH gradient, all wells were kept at pH 7 using Tris.HCl buffer, the pH gradient was also achieved with a Tris.HCl buffer.
The 96 well plates were then incubated while shaking while shaking for 2 hours at 37 degrees to allow time for GFP to mature. A longer incubation time was introduced to give more conclusive results. </p>
Triton X-100

Fig.1: Testing the effect of the detergent Triton X-100 on the CpxR promoter
Between the concentration values of 0.1-9% triton X-100 the data shows roughly the relationship that was expected, a rise in GFP production as the triton concentration increases. This is likely due to triton acting as a detergent resulting in damage to the plasma membrane of cells, activating the Cpx pathway in general, including the cpxR promoter. Past 10% Triton X-100 concentration it is assumed that cells are dying in large quantities, being unable to produce GFP at all or in very limited amounts before they are killed. This explains the corrected fluorescence readings reducing to below zero. The reason for the large spike in fluorescence at Triton concentrations >20% is unclear, a possibly explanation for this is the total lysis of the cell, expelling all cellular components resulting in background GFP levels having a larger fluorescence reading due to reduced scattering from within the cell
pH Effects
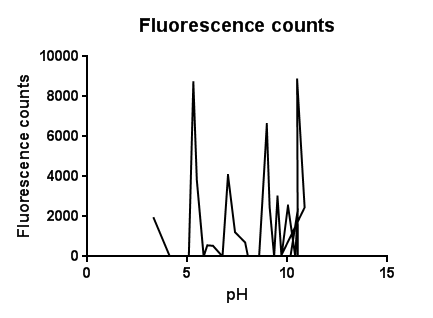
Fig.3: The effects of a range of pH values on the activation of the CpxR promoter
The slight fluctuations across the pH scale are in line with the background fluorescence readings from our cell cultures, representing the standard levels of cpx activation as the cells collide, disperse in solution and divide during replication.
The higher levels of expression at both low and high pH are likely to be due to the pH extremes denaturing proteins and disrupting interactions within the plasma membrane. It could be assumed that pH’s further into the extremes of acid and base would completely lyse and kill the cells resulting in a possibly lower fluorescence.
Atomic Force Microscopy

Fig.1: An image gained from Atomic Force Microscopy Experiments showing expression cells containing the pCpxR promoter coupled with GFP coding region.
The Atomic Force Microscopy experiments were designed to quantify the force necessary to trigger the CpxR promoter, generating GFP. This was to be done by observing the culture under the fluoroscope portion of the F-AFM, and then "prod" a bacterium at a low force, slowly increasing the stress until a fluorescence response was seen. This would then give a direct measure of the stress threshold for pCpxR, a value currently unknown in the literature. However, as with all new experiments, there were some teething issues. Firstly, it can be seen from the image that the culture fluoresced at a low level prior to exposure to the AFM tip. This was likely due to smaller stresses in solution and some can clearly be seen in the middle of binomial fission, which is known to also activate the Cpx pathway in general. There were also issues with trying to keep the bacterial cells stable enough to continually poke them with the AFM tip, initially a high glycerol concentration was employed in an attempt to achieve this however the cells remained very mobile.
Sequence and Features
- 10COMPATIBLE WITH RFC[10]
- 12COMPATIBLE WITH RFC[12]
- 21COMPATIBLE WITH RFC[21]
- 23COMPATIBLE WITH RFC[23]
- 25COMPATIBLE WITH RFC[25]
- 1000INCOMPATIBLE WITH RFC[1000]Illegal BsaI.rc site found at 728