As we have already clarified in the description part, self replicating shuttle vectors are essential for many
+
workflows, as the gene expression levels are higher and non of the tedious selection processes that come with
+
genomic integrations have to be done. <br>
+
+
On our road to the modular vector we were seeking, we firstly cured our own S. elongatus UTEX 2973 strain of its
+
pANS plasmid. This was done by transforming the pAM4787 vector, which holds a spectinomycin resistance as well as a
+
YFP cassette
+
<a href= https://www.microbiologyresearch.org/content/journal/micro/10.1099/mic.0.000377> (Chen et al., 2016)</a>.
+
Due to plasmid incompatibility - explained here in our design section <b>[Link to shuttle vector design]</b> - and because antibiotic pressure is applied, the pANS plasmid was over time cured from the
+
strain, which then just kept the pAM4787 plasmid. Transformation was done by conjugation with the pRK2013 plasmid in
+
DH5ɑ and the pAM4787 in HB101. Both were grown to an OD600≈0.5, washed in LB and mixed with S. elongatus which was
+
grown to late exponential phase and then washed in BG11.
+
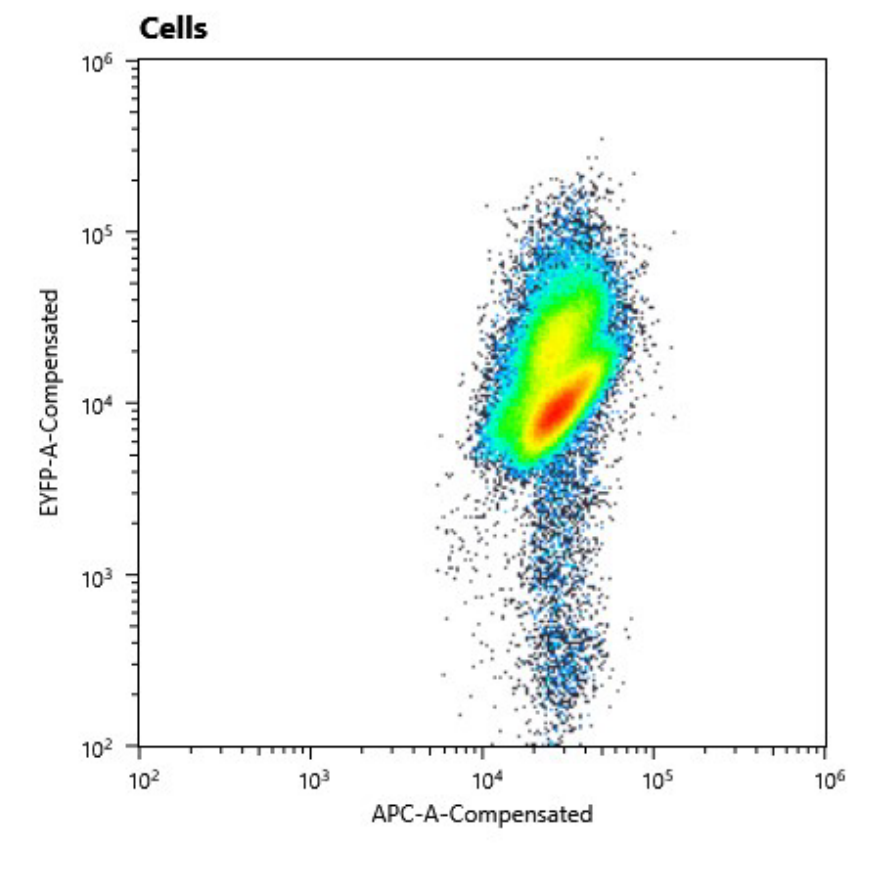
We could clearly show, that the conjugant strain bears the pAM4787 plasmid if selective pressure is held up.
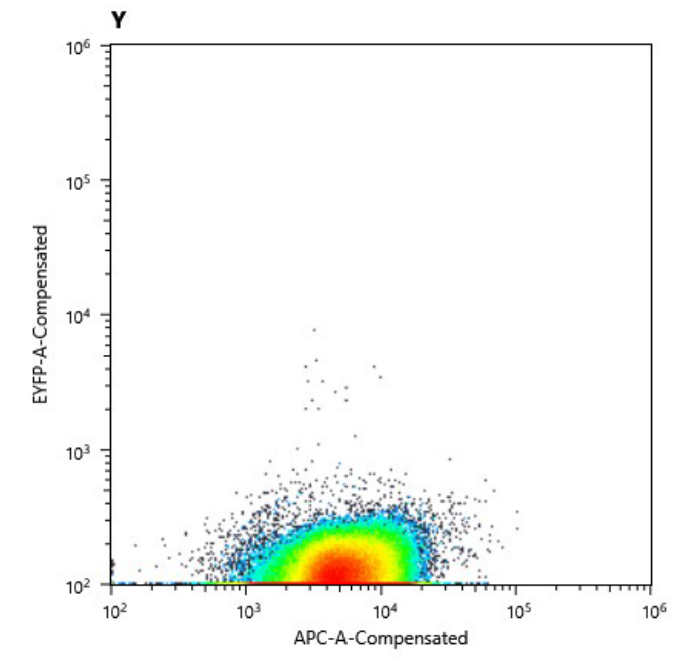
a) Cell counts of cured strain. The y axis shows relative YFP fluorescence and the x axis relative autofluorescence
+
b) Comparison of the fluorescence signal of the transformed (left) and cured (right) strain.
+
</figcaption> </figure>
+
+
+
+
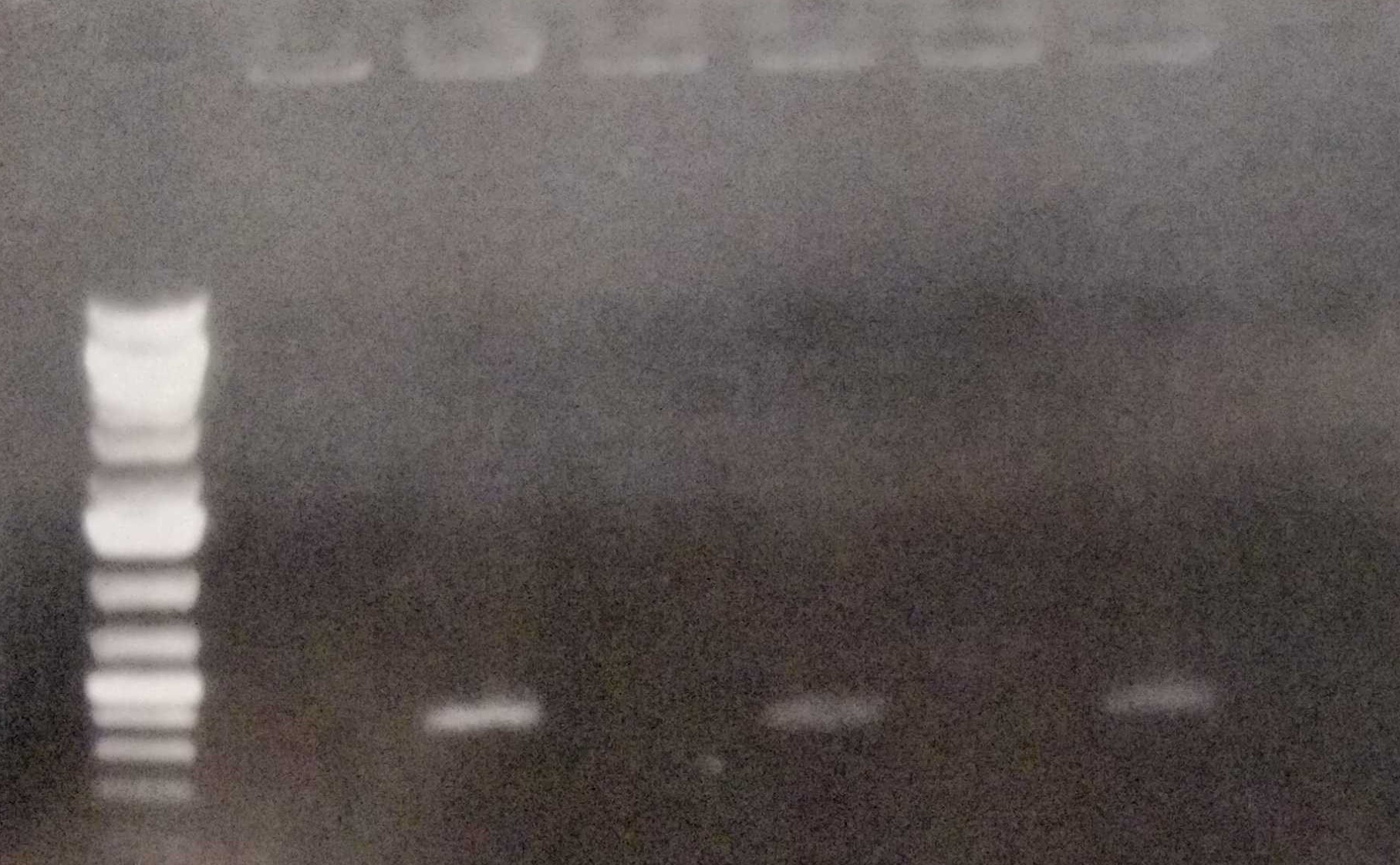
Furthermore we performed colony PCRs as a test. We sent our plasmid-free strain to Next Generation Sequencing in order to ensure that the strain really has lost the
<figcaption> Fig. 12: Colony PCR of the wild type, the conjugated and the cured strain.
+
</figcaption>
+
</figure>
+
+
Our next step was the characterization of the cyanobacterial shuttle vector mentioned in our design section <b>[Link to
+
design]</b>.
+
In an extensive flow cytometry experiment we assessed the fluorescence of a transformed YFP-construct in our cured
+
strain, showing that the shuttle vector with the minimal replication element can be maintained in<i>S. elongatus </i> UTEX
+
2973 .
+
<figure Style="text-align:center">
+
<img style="height: 65ex; width: 50ex"
+
src=
+
alt="hier kommen noch 3 bilder von FACS Messung aber no time">
+
<figcaption> Fig. 13: hier kommen noch 3 bilder von FACS Messung aber no time.
+
</figcaption>
+
</figure>
+
After another four weeks of cultivation we looked at our
+
cultures again on the UV table to check if fluorescence was still present and the high intensity of the fluorescence
+
proved to us, that the plasmid is still stably replicated in our strain, showing us, that the minimal replication
+
element does indeed work in our strain.
+
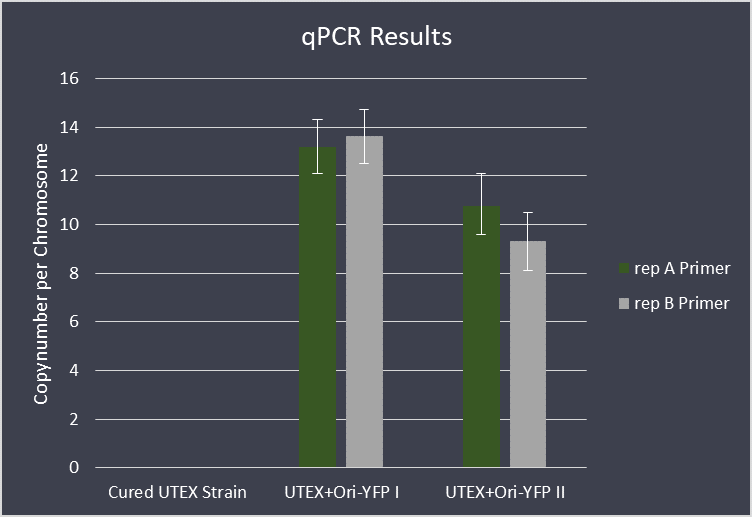
For further analysis we performed qPCR with this transformed strain, in order to check the copy number of the
+
vector. We used the copy number of pANL as a reference, which is supposedly at ~2,6 copies per chromosome
+
<a href= https://www.microbiologyresearch.org/content/journal/micro/10.1099/mic.0.000377> (Chen et al., 2016)</a>. Our data shows a ~4,5 times higher copy number relative to pANL, meaning that the construct is
+
maintained with approximately 11,7 copies per chromosome.
<figcaption> Fig. 15:YFP fluorescence at different optical densities. </figcaption>
+
</figure>
+
+
+
All this data confirms that the construct actually works and can be reliably used as a cyanobacterial shuttle
+
vector, proving that BBa_K3228069 works as intended, thus functioning as our validated part.
+
This assumption is solidified by all our sequence data, showing that the shuttle vectors were completely assembled
+
as planned in our design section <b>[Link to design of shuttle vectors]</b>
+
.
+
<figure Style="text-align:center">
+
<img style="height: 65ex; width: 50ex"
+
src= xyz
+
alt="disgramm">
+
<figcaption> Fig. 16: [FigXX seq results of lvl1 and lvl2 ori] </figcaption>
+
</figure>
+
</p>
+
+
</p>
+
</html>
<html>
<html>
<p align="justify">
<p align="justify">
−
+
This part is contained in the Green Expansion, a range of parts from <a href="https://2019.igem.org/Team:Marburg">iGEM Marburg 2019</a>that enables users of the Marburg Collection 2.0 to design MoClo compatible vectors for cyanobacteria as well as to engineer the genome of several cyanobacterial species.
−
This part is contained in the green expansion, a range of parts from iGEM Marburg 2020 that enables user of the Marburg Collection 2.0 to design MoClo compatible vectors for cyanobacteria as well as to engineer the genome of several cyanobacterial species.
+
</p>
</p>
</html>
</html>
−
===Overview===
+
===The Green Expansion===
<html>
<html>
<p align="justify">
<p align="justify">
−
The green expansion in the Marburg Collection 2.0 features the world's first MoClo compatible shuttle vector for cyanobacteria (BBa_K3228069). This composite part contains the origins of replication ColE1 (for cloning) and the ori of panS (for maintenance in cyanobacteria) as well as a spectinomycin cassette. It can be used in the Marburg Collection like any other composite part of the type 7+8 (antibiotic cassette + origin of replication).
+
The Green Expansion is an addition of parts to the Marburg Collection 2.0 <a href="http://2018.igem.org/Team:Marburg/Design">(See: Design of the Marburg Collection)</a> that features the world's first MoClo compatible shuttle vector for cyanobacteria. <a href="https://parts.igem.org/Part:BBa_K3228069">BBa_K3228069</a>
[[File:GE LVL1 ori.png|500px|thumb|left|'''Figure 1''': Design of the first MoClo compatible shuttle vector for cyanobacteria for LVL 1 constructs. This can be used for the integration of simple genetic modules. ]]
[[File:GE LVL2 ori.png|500px|thumb|left|'''Figure 2''': Design of the first MoClo compatible shuttle vector for cyanobacteria for LVL 2 constructs. This can be used for the integration of complex genetic devices. ]]
+
+
</div style>
+
+
<html>
+
The Green Expansion also offers all the parts needed for the genomic integration of one or multiple genes in cyanobacteria. This M.E.G.A. (Modularized Engineering of Genome Areas) kit convinces with a striking flexibility and a very intuitive workflow for the de novo assembly of your plasmid of choice. It encompasses five different neutral integration sites to choose from: three conventional sites frequently used in the cyanobacterial community (NSI to NSIII) as well as our own rationally designed artificial Neutral integration Site options a.N.S.o. 1 and 2 <a href="https://2019.igem.org/Team:Marburg/Design">(See: Finding new artificial Neutral integration Site options).</a>These sites show no transcriptional activity from neighboring regions according to RNA-seq data and are therefore completely orthogonal. Additionally we offer four different antibiotic markers to use (chloramphenicol, gentamicin, spectinomycin and kanamycin). With the Green Expansion up to 20 genes can be introduced into a cyanobacterial strain.
[[File:GE genomic integration.png|500px|thumb|left|'''Figure 3''': Overview over the modularized editing of genome area kit (M.E.G.A. kit) ]]
+
+
</div style>
+
<html>
<br>
<br>
<br>
<br>
−
This expansion also contains all the parts needed for the genomic integration of one or multiple genes in cyanobacteria. It convinces with a striking flexibility and a very intuitive workflow for the de novo assembly of your plasmid of choice. It encompasses 5 different neutral integration sites to choose from: three conventional sites frequently used in the cyanobacterial community (NSI to NSIII) as well as our own rationally designed artificial neutral integration site options (a.N.S.o. 1 and 2). These sites show no transcriptional activity from neighboring regions and are therefore completely orthogonal. Additionally we offer 4 different antibiotic markers to use (chloramphenicol, gentamycin, spectinomycin and kanamycin). In theory with this setup up to 20 genes could be introduced into a cyanobacterial strain.
+
Thanks to the flexible design this expansion can also be used for the genomic modification of any chassis after the introduction of a new species specific LVL 0 integration sites to our Marburg Collection 2.0. As the workflow to build new homologies is a bit more intricate compared to the one pot on step assembly of our other parts due to the internal BsmBI cutting site, we described the workflow for that in our design section <a href="https://2019.igem.org/Team:Marburg/Design">(See: Design of neutral integration sites).</a>
−
Thanks to the flexible design this expansion can also be used for the genomic modification of any chassis after the introduction of new species specific LVL 0 integration sites to our Marburg Collection 2.0. As the workflow to build new homologies is a bit more intricate compared to the one pot on step assembly of our other parts due to the internal BsmbI cutting site, we described the workflow for that in our design section (See Design: Design of the A.N.S.O.s).
+
<br>
<br>
<br>
<br>
−
The green expansion proves a valuable addition to our Marburg Collection 2.0 and to the iGEM Registry of Parts. It services users of our chassis and other cyanobacterial strains with a useful tool for genomic modifications but it also contributes a shell that can be used to modify any other model organism as well.
+
The Green Expansion proves a valuable addition to our Marburg Collection 2.0 and to the iGEM Registry of Standard Biological Parts. It services users of our chassis and other cyanobacterial strains with a useful tool for genomic modifications but it also contributes a shell that can be used to modify any other model organism as well.
+
</p>
+
</html>
+
+
===Compability===
+
<html>
+
<p align="justify">
+
+
These parts are compatible with the RCF [1000] standard and can be used in any part collection that uses the PhytoBrick standard of overhangs. For more information we recommend to head over to <a href="http://2018.igem.org/Team:Marburg/Design">Design of the Marburg Collection iGEM Marburg 2018</a>.
+
+
</p>
+
</html>
+
+
+
===iGEM Freiburg 2023: Testing and Documentation of BioBrick BBa_K3228069===
+
+
To modify <i>S. elongatus</i> PCC 7942, we, iGEM Freiburg 2023, decided to use this cyanobacteria-specific shuttle vector developed by iGEM Marburg 2019 which they kindly shared with us. This shuttle vector comes with 2 origins of replication (Ori): an Ori from PCC 7942, more specifically, from one of its endogenous pANS plasmids (which makes the vector compatible with our strain), and a high copy number Ori from <i>E. coli</i>, ColE1 (for more information, visit iGEM Marburg 2019 page.)
+
+
First, we cloned the aforementioned genes for B12 production (bluB and ssuE) into the shuttle vector, creating a new plasmid, piG_CBM. Next, we attempted to modify PCC 7942 with the piG_CBM via electroporation, conjugation (tri-parental mating), and natural transformation- none of which were successful (we observed no colonies on the plates containing antibiotic resistance). The reason for conjugation not succeeding was eventually found: the shuttle vector does not encompass an OriT (basis of mobility region/bom site) which would need to be added for the conjugation. Furthermore, according to Encinas et al. 2014 [1], the conjugative plasmid used as a helper and the shuttle vector should have the same OriT to improve the efficiency of conjugation.
+
+
The electroporation should not be affected by the lack of OriT and yet yielded no colonies after several attempts (with the exception of a faint colony for <i>S. sp.</i> PCC 6803. We did not find an explanation for why it is not working since the protocol we used from Prof Hess group, leader of the CyanoLab [2] at the University of Freiburg, was said to have a high success rate (at least for PCC 6803).
+
+
Possibly, a different, PCC 7942-specific electroporation protocol could be tried to further validate the shuttle vector. Also, the natural transformation did not succeed, however, we only tried it once (due to the time limitations) and this method of transformation has a lower efficiency, as a cyanobacteria-focused research group leader Prof. Wilde, also from the University of Freiburg [3], mentioned to us.
+
+
Therefore, several repetitions and/or a different transformation protocol might be needed to validate the shuttle vector’s applicability for natural transformation.
+
+
+
====iGEM Freiburg 2023: References====
+
+
[1]Encinas D, Garcillán-Barcia MP, Santos-Merino M, Delaye L, Moya A, De La Cruz F. Plasmid Conjugation from Proteobacteria as Evidence for the Origin of Xenologous Genes in Cyanobacteria. Journal of Bacteriology [Internet]. 2014 Apr 15;196(8):1551–9. Available from: https://doi.org/10.1128/jb.01464-13
Illegal XbaI site found at 1199 Illegal PstI site found at 3734
12
INCOMPATIBLE WITH RFC[12]
Illegal PstI site found at 3734 Illegal NotI site found at 1 Illegal NotI site found at 5738
21
INCOMPATIBLE WITH RFC[21]
Illegal BglII site found at 4897 Illegal XhoI site found at 1249
23
INCOMPATIBLE WITH RFC[23]
Illegal XbaI site found at 1199 Illegal PstI site found at 3734
25
INCOMPATIBLE WITH RFC[25]
Illegal XbaI site found at 1199 Illegal PstI site found at 3734 Illegal NgoMIV site found at 48 Illegal NgoMIV site found at 190
1000
COMPATIBLE WITH RFC[1000]
B A S I C P A R T S
Cyanobacterial shuttle vectors
As we have already clarified in the description part, self replicating shuttle vectors are essential for many
workflows, as the gene expression levels are higher and non of the tedious selection processes that come with
genomic integrations have to be done.
On our road to the modular vector we were seeking, we firstly cured our own S. elongatus UTEX 2973 strain of its
pANS plasmid. This was done by transforming the pAM4787 vector, which holds a spectinomycin resistance as well as a
YFP cassette
(Chen et al., 2016).
Due to plasmid incompatibility - explained here in our design section [Link to shuttle vector design] - and because antibiotic pressure is applied, the pANS plasmid was over time cured from the
strain, which then just kept the pAM4787 plasmid. Transformation was done by conjugation with the pRK2013 plasmid in
DH5ɑ and the pAM4787 in HB101. Both were grown to an OD600≈0.5, washed in LB and mixed with S. elongatus which was
grown to late exponential phase and then washed in BG11.
We could clearly show, that the conjugant strain bears the pAM4787 plasmid if selective pressure is held up.
Fig. 11: Cell counts of conjugant strain. The y axis shows relative YFP fluorescence and the x axis relative autofluorescence.
This was followed by us starting to culture the pAM4787 bearing strain without
antibiotics again, slowly removing selective pressure from the cells. As the plasmid does not give them any other
advantage and is probably just more metabolic burden due to the constantly produced YFP proteins it is slowly being
lost.
We could prove this in multiple setups: with the flow cytometry device we were kindly granted access to we could
clearly show the missing YFP signal in the cured S. elongatus strain and
logically this could also be observed over our UV table
a) Cell counts of cured strain. The y axis shows relative YFP fluorescence and the x axis relative autofluorescence
b) Comparison of the fluorescence signal of the transformed (left) and cured (right) strain.
Furthermore we performed colony PCRs as a test. We sent our plasmid-free strain to Next Generation Sequencing in order to ensure that the strain really has lost the
pANS plasmid.
Fig. 12: Colony PCR of the wild type, the conjugated and the cured strain.
Our next step was the characterization of the cyanobacterial shuttle vector mentioned in our design section [Link to
design].
In an extensive flow cytometry experiment we assessed the fluorescence of a transformed YFP-construct in our cured
strain, showing that the shuttle vector with the minimal replication element can be maintained inS. elongatus UTEX
2973 .
Fig. 13: hier kommen noch 3 bilder von FACS Messung aber no time.
After another four weeks of cultivation we looked at our
cultures again on the UV table to check if fluorescence was still present and the high intensity of the fluorescence
proved to us, that the plasmid is still stably replicated in our strain, showing us, that the minimal replication
element does indeed work in our strain.
For further analysis we performed qPCR with this transformed strain, in order to check the copy number of the
vector. We used the copy number of pANL as a reference, which is supposedly at ~2,6 copies per chromosome
(Chen et al., 2016). Our data shows a ~4,5 times higher copy number relative to pANL, meaning that the construct is
maintained with approximately 11,7 copies per chromosome.
Fig. 14: Copynumber Evaluation of Ori-part via qPCR
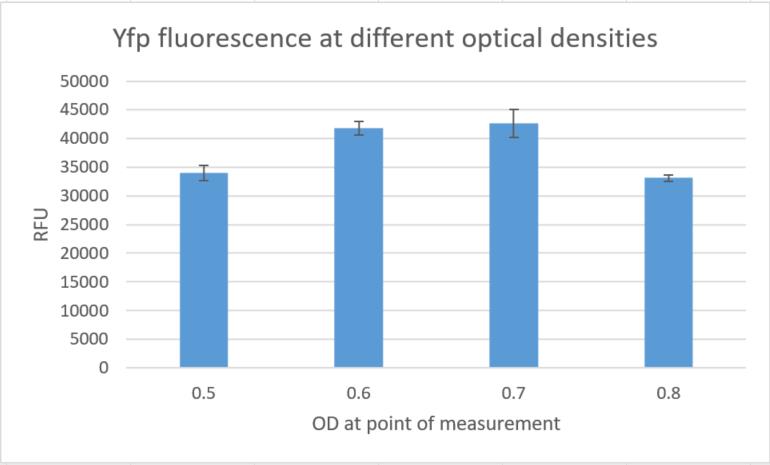
Additionally we measured the fluorescence signals in a plate reader at different optical densities and could again
confirm high fluorescence signals, indicating strong gene expression in constructs built around this replication
element.
Fig. 15:YFP fluorescence at different optical densities.
All this data confirms that the construct actually works and can be reliably used as a cyanobacterial shuttle
vector, proving that BBa_K3228069 works as intended, thus functioning as our validated part.
This assumption is solidified by all our sequence data, showing that the shuttle vectors were completely assembled
as planned in our design section [Link to design of shuttle vectors]
.
Fig. 16: [FigXX seq results of lvl1 and lvl2 ori]
This part is contained in the Green Expansion, a range of parts from iGEM Marburg 2019that enables users of the Marburg Collection 2.0 to design MoClo compatible vectors for cyanobacteria as well as to engineer the genome of several cyanobacterial species.
The Green Expansion
The Green Expansion is an addition of parts to the Marburg Collection 2.0 (See: Design of the Marburg Collection) that features the world's first MoClo compatible shuttle vector for cyanobacteria. BBa_K3228069
Figure 1: Design of the first MoClo compatible shuttle vector for cyanobacteria for LVL 1 constructs. This can be used for the integration of simple genetic modules.
Figure 2: Design of the first MoClo compatible shuttle vector for cyanobacteria for LVL 2 constructs. This can be used for the integration of complex genetic devices.
The Green Expansion also offers all the parts needed for the genomic integration of one or multiple genes in cyanobacteria. This M.E.G.A. (Modularized Engineering of Genome Areas) kit convinces with a striking flexibility and a very intuitive workflow for the de novo assembly of your plasmid of choice. It encompasses five different neutral integration sites to choose from: three conventional sites frequently used in the cyanobacterial community (NSI to NSIII) as well as our own rationally designed artificial Neutral integration Site options a.N.S.o. 1 and 2 (See: Finding new artificial Neutral integration Site options).These sites show no transcriptional activity from neighboring regions according to RNA-seq data and are therefore completely orthogonal. Additionally we offer four different antibiotic markers to use (chloramphenicol, gentamicin, spectinomycin and kanamycin). With the Green Expansion up to 20 genes can be introduced into a cyanobacterial strain.
Figure 3: Overview over the modularized editing of genome area kit (M.E.G.A. kit)
Thanks to the flexible design this expansion can also be used for the genomic modification of any chassis after the introduction of a new species specific LVL 0 integration sites to our Marburg Collection 2.0. As the workflow to build new homologies is a bit more intricate compared to the one pot on step assembly of our other parts due to the internal BsmBI cutting site, we described the workflow for that in our design section (See: Design of neutral integration sites).
The Green Expansion proves a valuable addition to our Marburg Collection 2.0 and to the iGEM Registry of Standard Biological Parts. It services users of our chassis and other cyanobacterial strains with a useful tool for genomic modifications but it also contributes a shell that can be used to modify any other model organism as well.
Compability
These parts are compatible with the RCF [1000] standard and can be used in any part collection that uses the PhytoBrick standard of overhangs. For more information we recommend to head over to Design of the Marburg Collection iGEM Marburg 2018.
iGEM Freiburg 2023: Testing and Documentation of BioBrick BBa_K3228069
To modify S. elongatus PCC 7942, we, iGEM Freiburg 2023, decided to use this cyanobacteria-specific shuttle vector developed by iGEM Marburg 2019 which they kindly shared with us. This shuttle vector comes with 2 origins of replication (Ori): an Ori from PCC 7942, more specifically, from one of its endogenous pANS plasmids (which makes the vector compatible with our strain), and a high copy number Ori from E. coli, ColE1 (for more information, visit iGEM Marburg 2019 page.)
First, we cloned the aforementioned genes for B12 production (bluB and ssuE) into the shuttle vector, creating a new plasmid, piG_CBM. Next, we attempted to modify PCC 7942 with the piG_CBM via electroporation, conjugation (tri-parental mating), and natural transformation- none of which were successful (we observed no colonies on the plates containing antibiotic resistance). The reason for conjugation not succeeding was eventually found: the shuttle vector does not encompass an OriT (basis of mobility region/bom site) which would need to be added for the conjugation. Furthermore, according to Encinas et al. 2014 [1], the conjugative plasmid used as a helper and the shuttle vector should have the same OriT to improve the efficiency of conjugation.
The electroporation should not be affected by the lack of OriT and yet yielded no colonies after several attempts (with the exception of a faint colony for S. sp. PCC 6803. We did not find an explanation for why it is not working since the protocol we used from Prof Hess group, leader of the CyanoLab [2] at the University of Freiburg, was said to have a high success rate (at least for PCC 6803).
Possibly, a different, PCC 7942-specific electroporation protocol could be tried to further validate the shuttle vector. Also, the natural transformation did not succeed, however, we only tried it once (due to the time limitations) and this method of transformation has a lower efficiency, as a cyanobacteria-focused research group leader Prof. Wilde, also from the University of Freiburg [3], mentioned to us.
Therefore, several repetitions and/or a different transformation protocol might be needed to validate the shuttle vector’s applicability for natural transformation.
iGEM Freiburg 2023: References
[1]Encinas D, Garcillán-Barcia MP, Santos-Merino M, Delaye L, Moya A, De La Cruz F. Plasmid Conjugation from Proteobacteria as Evidence for the Origin of Xenologous Genes in Cyanobacteria. Journal of Bacteriology [Internet]. 2014 Apr 15;196(8):1551–9. Available from: https://doi.org/10.1128/jb.01464-13